

TMEM132A regulates Wnt/β-catenin signaling through stabilizing LRP6 during mouse embryonic development
- PMID: 39385148
- PMCID: PMC11465819
- DOI: 10.1186/s12964-024-01855-9
TMEM132A regulates Wnt/β-catenin signaling through stabilizing LRP6 during mouse embryonic development
Abstract
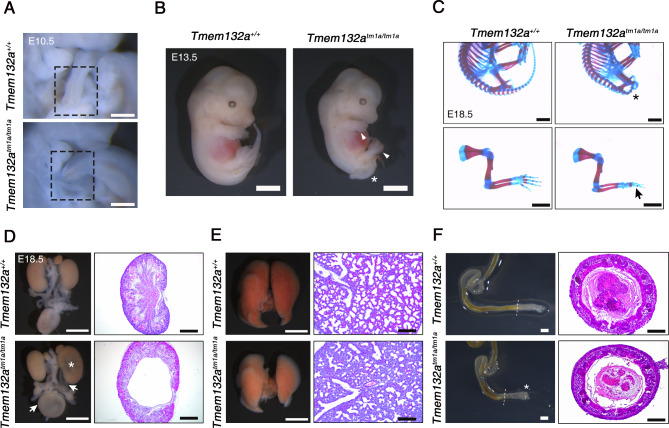
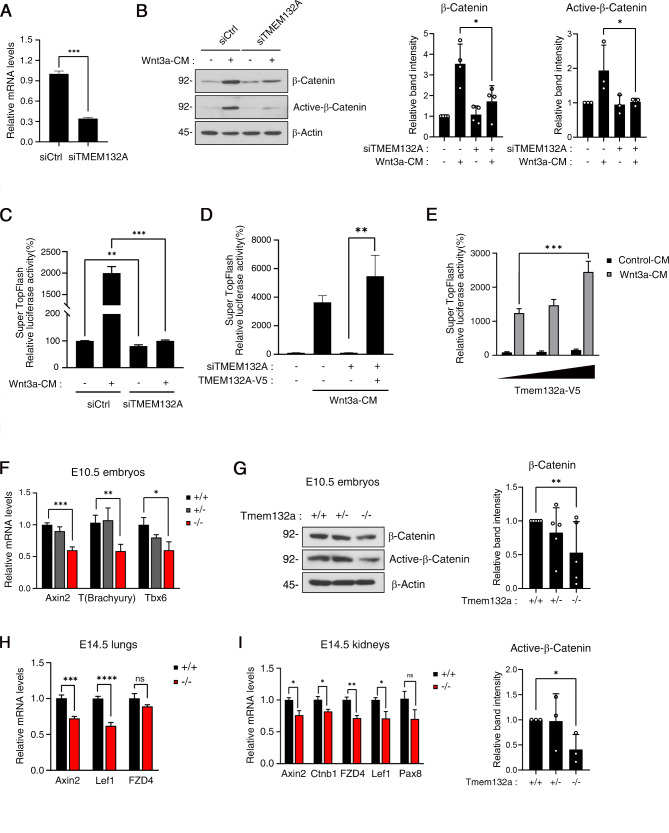
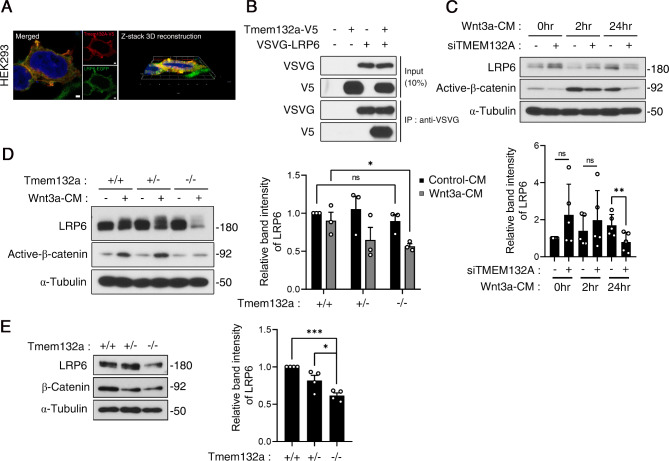
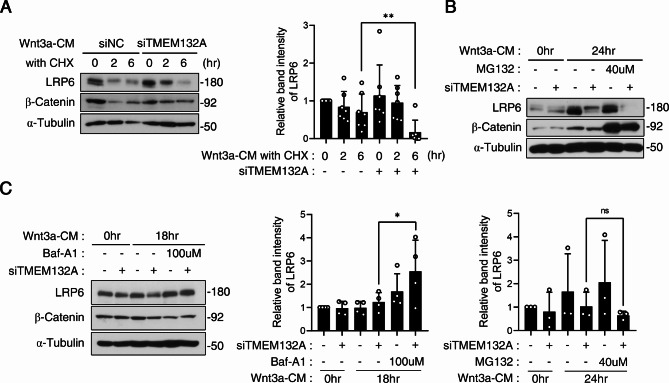
The Wnt/β-catenin signaling pathway is crucial for embryonic development and adult tissue homeostasis. Dysregulation of Wnt signaling is linked to various developmental anomalies and diseases, notably cancer. Although numerous regulators of the Wnt signaling pathway have been identified, their precise function during mouse embryo development remains unclear. Here, we revealed that TMEM132A is a crucial regulator of canonical Wnt/β-catenin signaling in mouse development. Mouse embryos lacking Tmem132a displayed a range of malformations, including open spina bifida, caudal truncation, syndactyly, and renal defects, similar to the phenotypes of Wnt/β-catenin mutants. Tmem132a knockdown in cultured cells suppressed canonical Wnt/β-catenin signaling. In developing mice, loss of Tmem132a also led to diminished Wnt/β-catenin signaling. Mechanistically, we showed that TMEM132A interacts with the Wnt co-receptor LRP6, thereby stabilizing it and preventing its lysosomal degradation. These findings shed light on a novel role for TMEM132A in regulating LRP6 stability and canonical Wnt/β-catenin signaling during mouse embryo development. This study provides valuable insights into the molecular intricacies of the Wnt signaling pathway. Further research may deepen our understanding of Wnt pathway regulation and offer its potential therapeutic applications.
Keywords: LRP6; Lysosomal degradation; Mouse development; TMEM132A; Wnt/β-catenin signaling.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
