

Thaumarchaeota from deep-sea methane seeps provide novel insights into their evolutionary history and ecological implications
- PMID: 39385283
- PMCID: PMC11463064
- DOI: 10.1186/s40168-024-01912-y
Thaumarchaeota from deep-sea methane seeps provide novel insights into their evolutionary history and ecological implications
Abstract
Background: Ammonia-oxidizing archaea (AOA) of the phylum Thaumarchaeota mediate the rate-limiting step of nitrification and remove the ammonia that inhibits the aerobic metabolism of methanotrophs. However, the AOA that inhabit deep-sea methane-seep surface sediments (DMS) are rarely studied. Here, we used global DMS metagenomics and metagenome-assembled genomes (MAGs) to investigate the metabolic activity, evolutionary history, and ecological contributions of AOA. Expression of AOA-specific ammonia-oxidizing gene (amoA) was examined in the sediments collected from the South China Sea (SCS) to identify their active ammonia metabolism in the DMS.
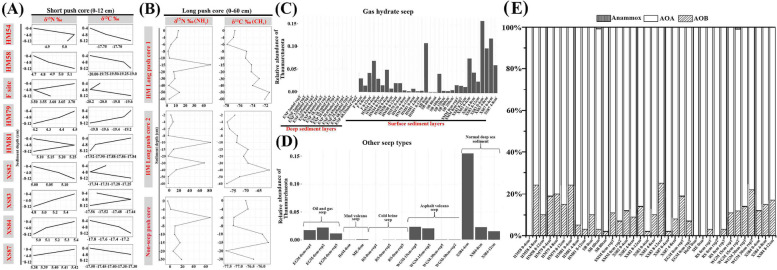
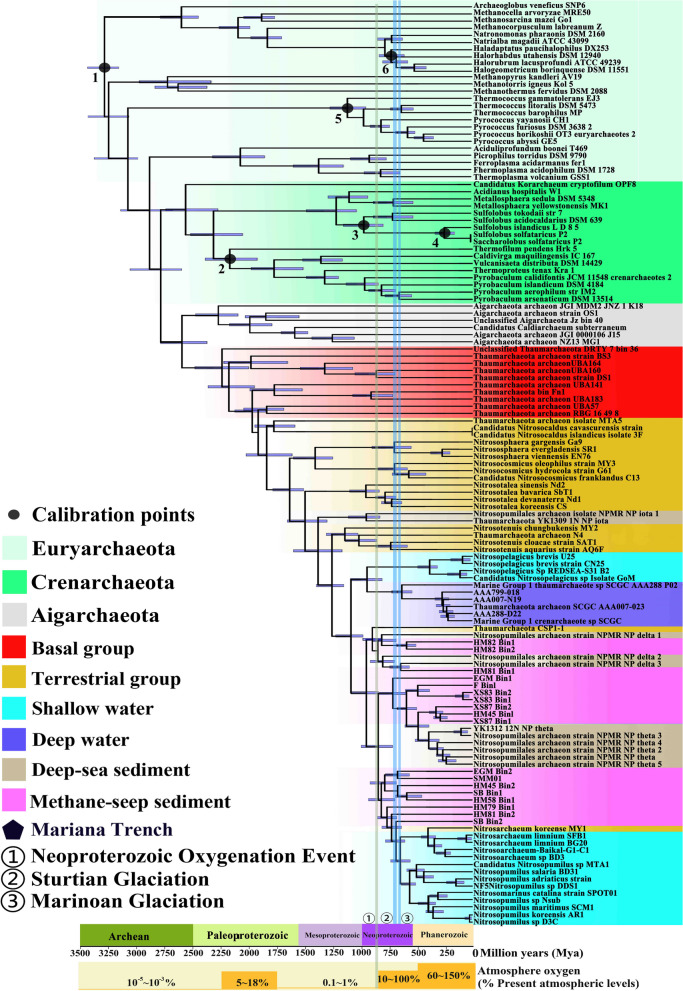
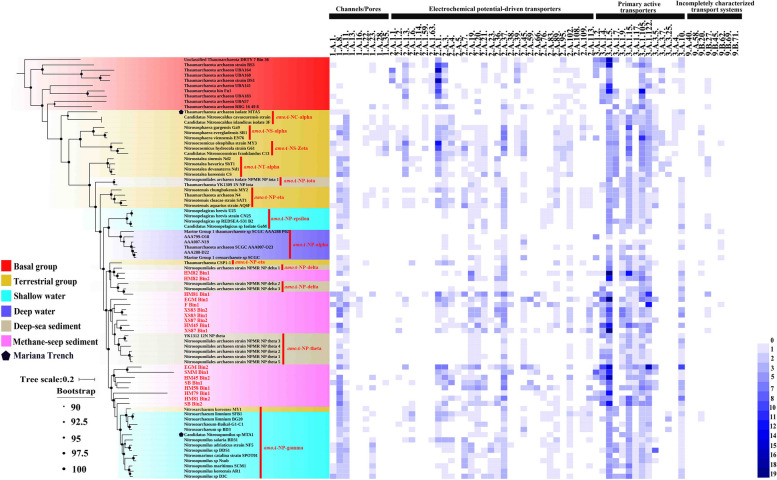
Results: Our analysis indicated that AOA contribute > 75% to the composition of ammonia-utilization genes within the surface layers (above 30 cm) of global DMS. The AOA-specific ammonia-oxidizing gene was actively expressed in the DMS collected from the SCS. Phylogenomic analysis of medium-/high-quality MAGs from 18 DMS-AOA indicated that they evolved from ancestors in the barren deep-sea sediment and then expanded from the DMS to shallow water forming an amoA-NP-gamma clade-affiliated lineage. Molecular dating suggests that the DMS-AOA origination coincided with the Neoproterozoic oxidation event (NOE), which occurred ~ 800 million years ago (mya), and their expansion to shallow water coincided with the Sturtian glaciation (~ 713 mya). Comparative genomic analysis suggests that DMS-AOA exhibit higher requirement of carbon source for protein synthesis with enhanced genomic capability for osmotic regulation, motility, chemotaxis, and utilization of exogenous organic compounds, suggesting it could be more heterotrophic compared with other lineages.
Conclusion: Our findings provide new insights into the evolutionary history of AOA within the Thaumarchaeota, highlighting their critical roles in nitrogen cycling in the global DMS ecosystems. Video Abstract.
Keywords: Ammonia-oxidizing archaea; Deep-sea sediment; Genomic evolution; Methane seep.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Jørgensen BB, Boetius A. Feast and famine—microbial life in the deep-sea bed. Nat Rev Microbiol. 2007;5(10):770–81. - PubMed
-
- Boetius A, Wenzhöfer F. Seafloor oxygen consumption fuelled by methane from cold seeps. Nat Geosci. 2013;6(9):725–34.
-
- Åström EK, et al. Methane cold seeps as biological oases in the high-Arctic deep sea. Limnol Oceanogr. 2018;63(S1):S209–31.
-
- Leininger S, et al. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature. 2006;442(7104):806–9. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
