

This is a preprint.
The formation and propagation of human Robertsonian chromosomes
- PMID: 39386535
- PMCID: PMC11463614
- DOI: 10.1101/2024.09.24.614821
The formation and propagation of human Robertsonian chromosomes
Update in
-
The formation and propagation of human Robertsonian chromosomes.Nature. 2025 Nov;647(8091):952-961. doi: 10.1038/s41586-025-09540-8. Epub 2025 Sep 24. Nature. 2025. PMID: 40993387 Free PMC article.
Abstract
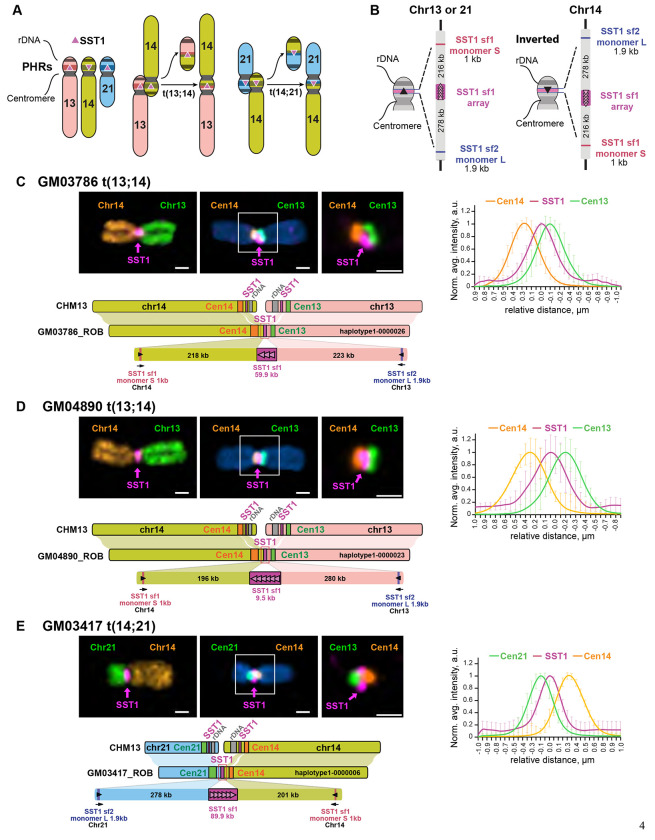
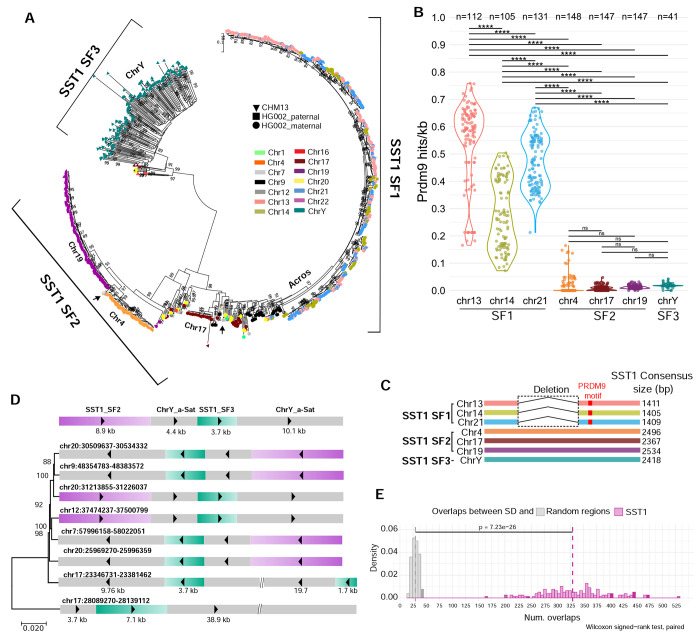
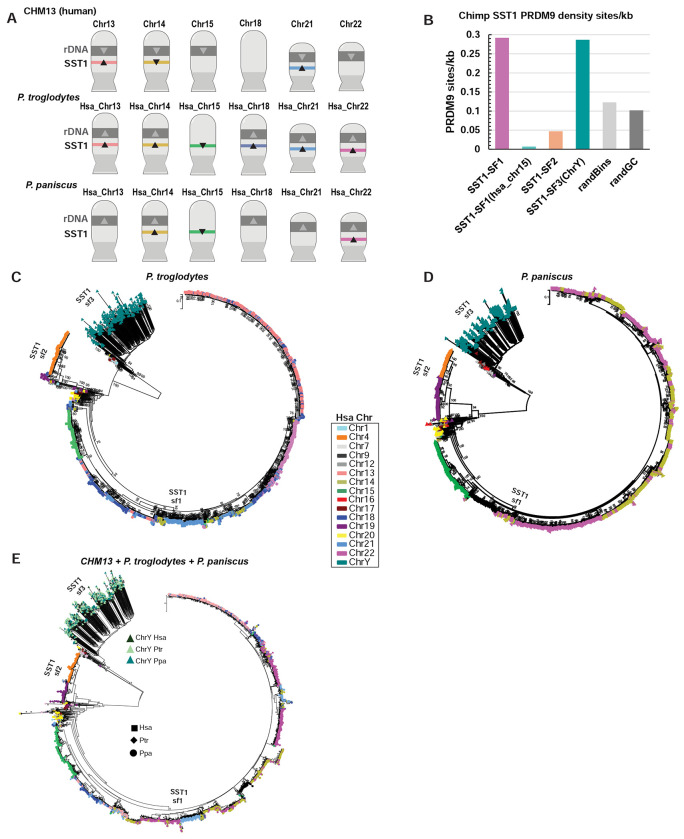
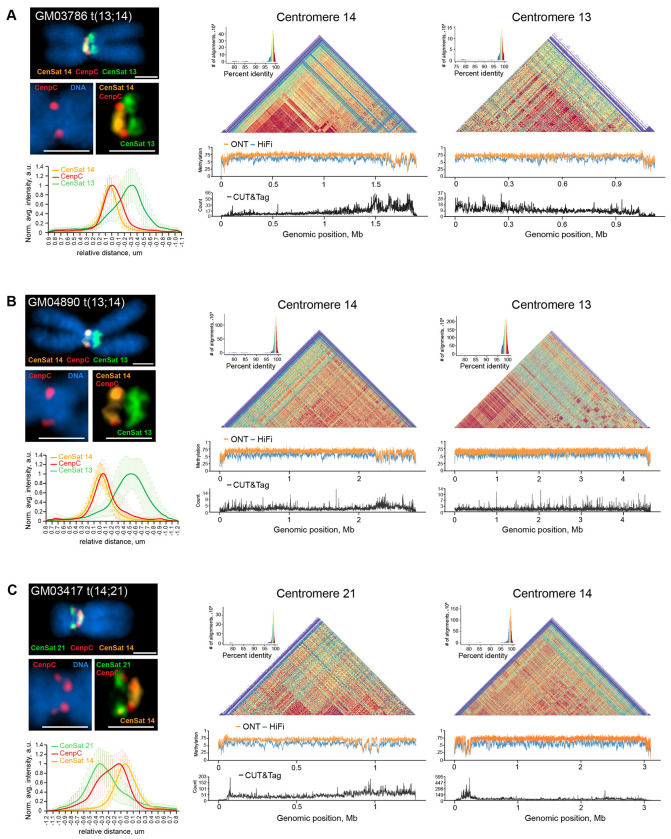
Robertsonian chromosomes are a type of variant chromosome found commonly in nature. Present in one in 800 humans, these chromosomes can underlie infertility, trisomies, and increased cancer incidence. Recognized cytogenetically for more than a century, their origins have remained mysterious. Recent advances in genomics allowed us to assemble three human Robertsonian chromosomes completely. We identify a common breakpoint and epigenetic changes in centromeres that provide insight into the formation and propagation of common Robertsonian translocations. Further investigation of the assembled genomes of chimpanzee and bonobo highlights the structural features of the human genome that uniquely enable the specific crossover event that creates these chromosomes. Resolving the structure and epigenetic features of human Robertsonian chromosomes at a molecular level paves the way to understanding how chromosomal structural variation occurs more generally, and how chromosomes evolve.
Figures
References
-
- Robertson. Chromosome studies. I. Taxonomic relationships shown in the chromosomes of Tettigidae and Acrididae: V-shaped chromosomes and their significant in Acridia, Locustidae, and Gryllidae: Chromosomes and variation. J Morphology 27, 179–331 (1916).
-
- Schubert I., Schriever-Schwemmer G., Werner T. & Adler I. D. Telomeric signals in robertsonian fusion and fission chromosomes: implications for the origin of pseudoaneuploidy. Cytogenetics and cell genetics 59, 6–9 (1992). - PubMed
-
- Glugoski L., Deon G. A., Nogaroto V., Moreira-Filho O. & Vicari M. R. Robertsonian fusion site in Rineloricaria pentamaculata (Siluriformes: Loricariidae): involvement of 5S rDNA and satellite sequences. Cytogenetic and genome research (2023). - PubMed
-
- Lukhtanov V. A. et al. Reinforcement of pre-zygotic isolation and karyotype evolution in Agrodiaetus butterflies. Nature 436, 385–389 (2005). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources