

This is a preprint.
Genomic mosaicism reveals developmental organization of trunk neural crest-derived ganglia
- PMID: 39386543
- PMCID: PMC11463384
- DOI: 10.1101/2024.09.25.615004
Genomic mosaicism reveals developmental organization of trunk neural crest-derived ganglia
Abstract
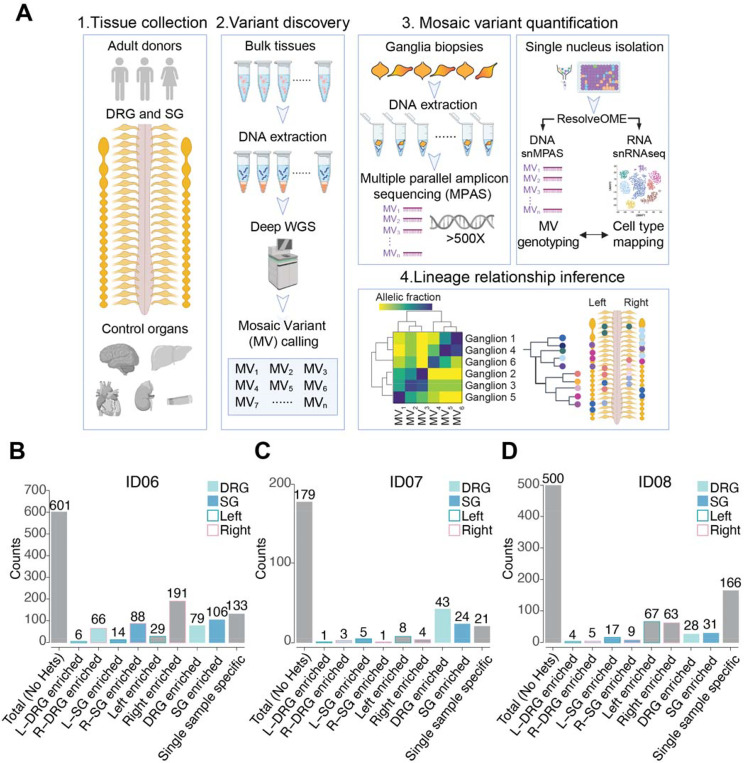
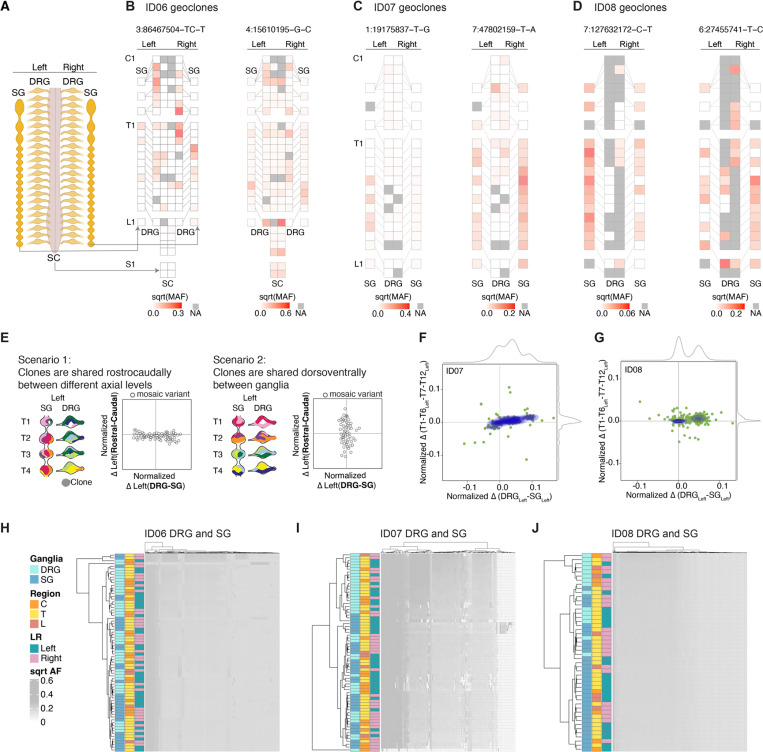
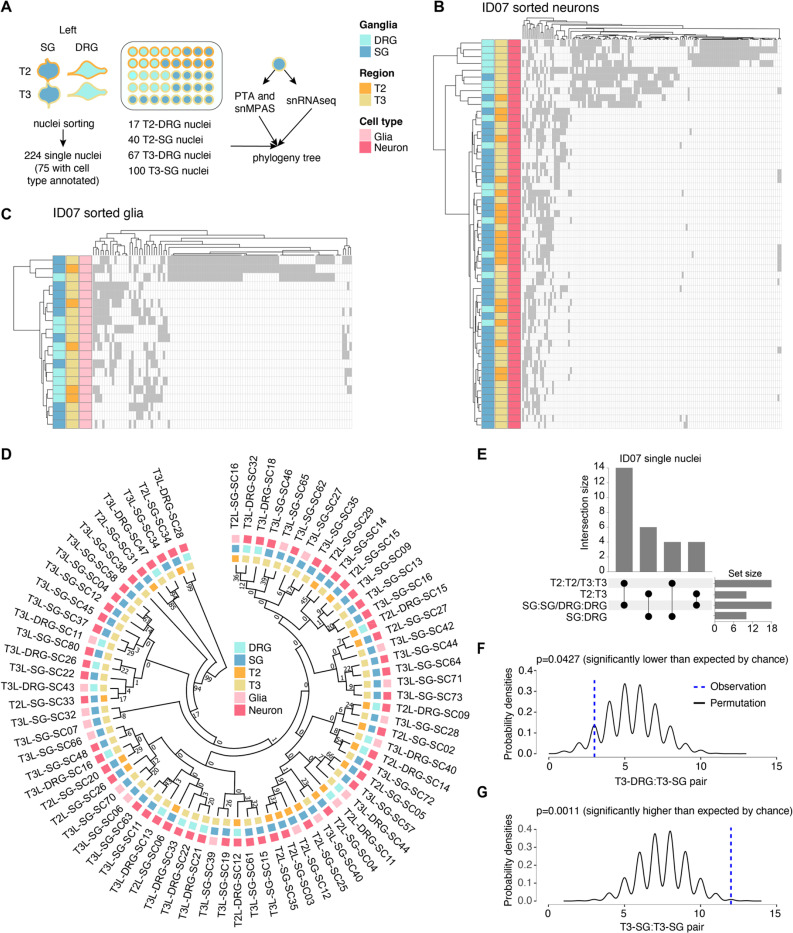
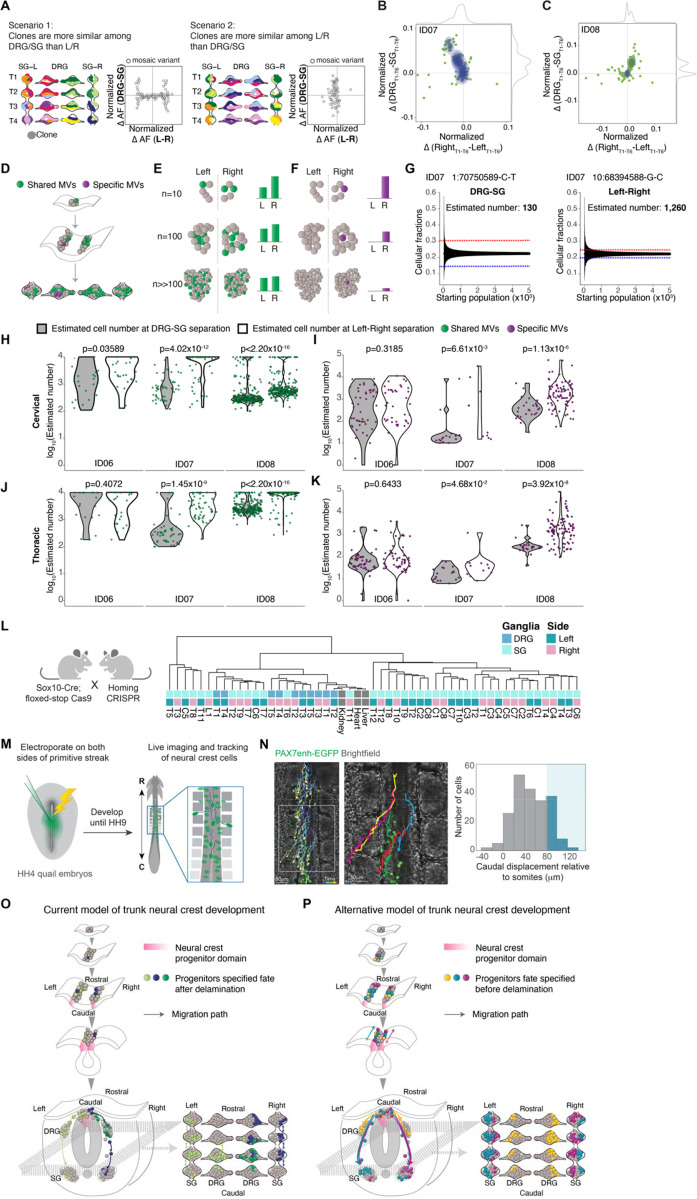
The neural crest generates numerous cell types, but conflicting results leave developmental origins unresolved. Here using somatic mosaic variants as cellular barcodes, we infer embryonic clonal dynamics of trunk neural crest, focusing on the sensory and sympathetic ganglia. From three independent adult neurotypical human donors, we identified 1,278 mosaic variants using deep whole-genome sequencing, then profiled allelic fractions in 187 anatomically dissected ganglia. We found a massive rostrocaudal spread of progenitor clones specific to sensory or sympathetic ganglia, which unlike in the brain, showed robust bilateral distributions. Computational modeling suggested neural crest progenitor fate specification preceded delamination from neural tube. Single-cell multiomic analysis suggested both neurons and glia contributed to the rostrocaudal clonal organization. CRISPR barcoding in mice and live imaging in quail embryos confirmed these clonal dynamics across multiple somite levels. Our findings reveal an evolutionarily conserved clonal spread of cells populating peripheral neural ganglia.
Keywords: cell migration; dorsal root ganglia; lineage tracing; live imaging; neural crest; somatic mosaicism; sympathetic ganglia.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources