

This is a preprint.
Functional redundancy between penicillin-binding proteins during asymmetric cell division in Clostridioides difficile
- PMID: 39386573
- PMCID: PMC11463367
- DOI: 10.1101/2024.09.26.615255
Functional redundancy between penicillin-binding proteins during asymmetric cell division in Clostridioides difficile
Update in
-
Penicillin-binding proteins exhibit functional redundancy during asymmetric cell division in Clostridioides difficile.J Bacteriol. 2025 Dec 18;207(12):e0050325. doi: 10.1128/jb.00503-25. Epub 2025 Nov 26. J Bacteriol. 2025. PMID: 41294337 Free PMC article.
Abstract
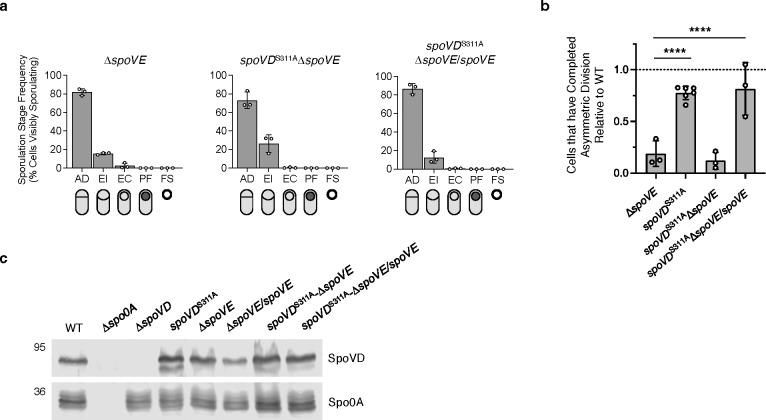
Peptidoglycan synthesis is an essential driver of bacterial growth and division. The final steps of this crucial process involve the activity of the SEDS family glycosyltransferases that polymerize glycan strands and the class B penicillin-binding protein (bPBP) transpeptidases that cross-link them. While many bacteria encode multiple bPBPs to perform specialized roles during specific cellular processes, some bPBPs can play redundant roles that are important for resistance against certain cell wall stresses. Our understanding of these compensatory mechanisms, however, remains incomplete. Endospore-forming bacteria typically encode multiple bPBPs that drive morphological changes required for sporulation. The sporulation-specific bPBP, SpoVD, is important for synthesizing the asymmetric division septum and spore cortex peptidoglycan during sporulation in the pathogen Clostridioides difficile. Although SpoVD catalytic activity is essential for cortex synthesis, we show that it is unexpectedly dispensable for SpoVD to mediate asymmetric division. The dispensability of SpoVD's catalytic activity requires the presence of its SEDS partner, SpoVE, and is facilitated by another sporulation-induced bPBP, PBP3. Our data further suggest that PBP3 interacts with components of the asymmetric division machinery, including SpoVD. These findings suggest a possible mechanism by which bPBPs can be functionally redundant in diverse bacteria and facilitate antibiotic resistance.
Figures





References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources