

This is a preprint.
A unique interplay of access and selection shapes peritoneal metastasis evolution in colorectal cancer
- PMID: 39386634
- PMCID: PMC11463674
- DOI: 10.1101/2024.09.25.614736
A unique interplay of access and selection shapes peritoneal metastasis evolution in colorectal cancer
Abstract
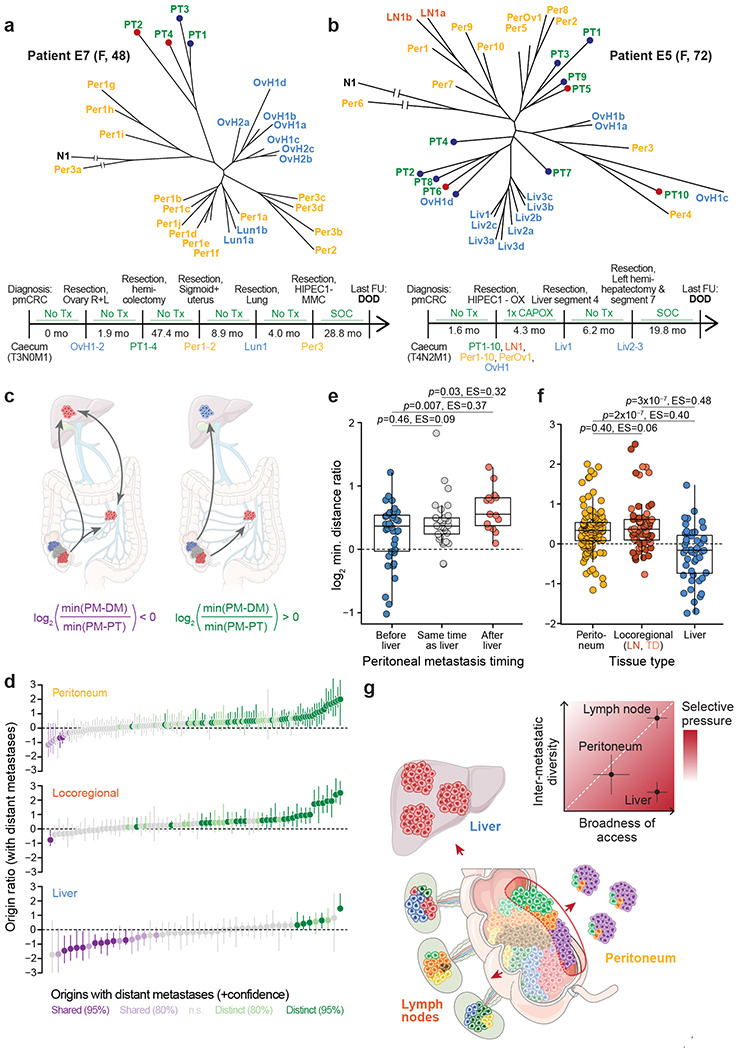
Whether metastasis in humans can be accomplished by most primary tumor cells or requires the evolution of a specialized trait remains an open question. To evaluate whether metastases are founded by non-random subsets of primary tumor lineages requires extensive, difficult-to-implement sampling. We have realized an unusually dense multi-region sampling scheme in a cohort of 26 colorectal cancer patients with peritoneal metastases, reconstructing the evolutionary history of on average 28.8 tissue samples per patient with a microsatellite-based fingerprinting assay. To assess metastatic randomness, we evaluate inter- and intra-metastatic heterogeneity relative to the primary tumor and find that peritoneal metastases are more heterogeneous than liver metastases but less diverse than locoregional metastases. Metachronous peritoneal metastases exposed to systemic chemotherapy show significantly higher inter-lesion diversity than synchronous, untreated metastases. Projection of peritoneal metastasis origins onto a spatial map of the primary tumor reveals that they often originate at the deep-invading edge, in contrast to liver and lymph node metastases which exhibit no such preference. Furthermore, peritoneal metastases typically do not share a common subclonal origin with distant metastases in more remote organs. Synthesizing these insights into an evolutionary portrait of peritoneal metastases, we conclude that the peritoneal-metastatic process imposes milder selective pressures onto disseminating cancer cells than the liver-metastatic process. Peritoneal metastases' unique evolutionary features have potential implications for staging and treatment.
Conflict of interest statement
Competing financial interests The authors declare no competing financial interests.
Figures




References
-
- McPherson A. et al. Divergent modes of clonal spread and intraperitoneal mixing in high-grade serous ovarian cancer. Nat. Genet. 48, 758–767 (2016). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources