

This is a preprint.
An atlas of RNA-dependent proteins in cell division reveals the riboregulation of mitotic protein-protein interactions
- PMID: 39386702
- PMCID: PMC11463612
- DOI: 10.1101/2024.09.25.614981
An atlas of RNA-dependent proteins in cell division reveals the riboregulation of mitotic protein-protein interactions
Update in
-
An atlas of RNA-dependent proteins in cell division reveals the riboregulation of mitotic protein-protein interactions.Nat Commun. 2025 Mar 8;16(1):2325. doi: 10.1038/s41467-025-57671-3. Nat Commun. 2025. PMID: 40057470 Free PMC article.
Abstract
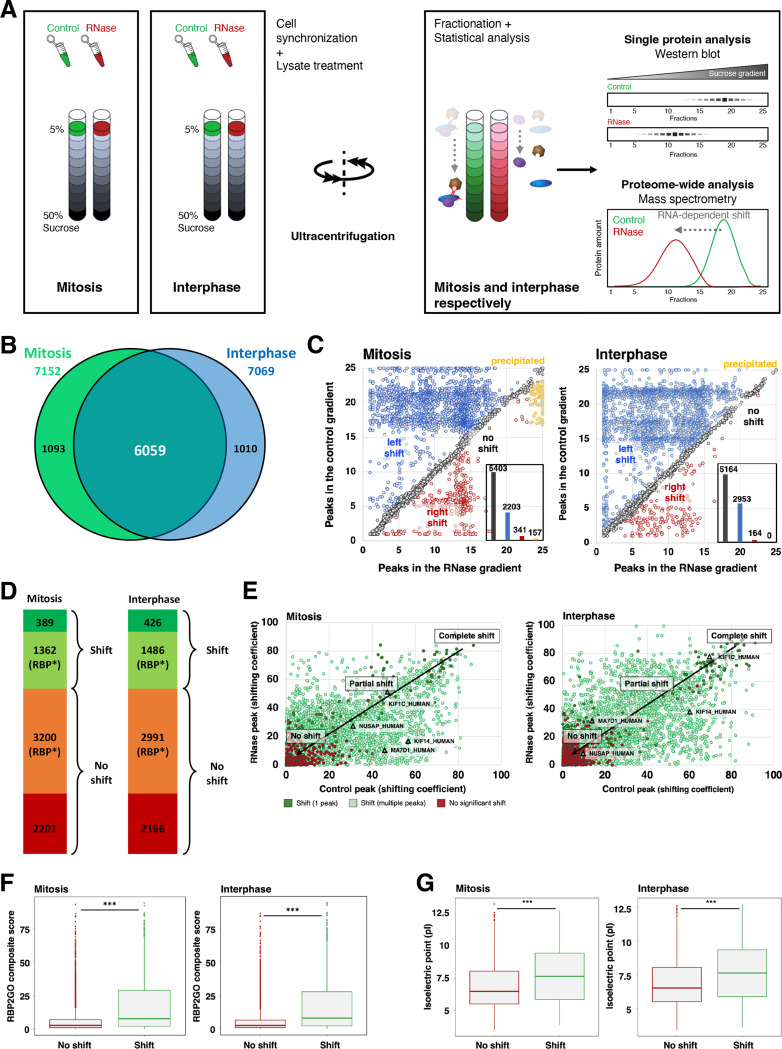
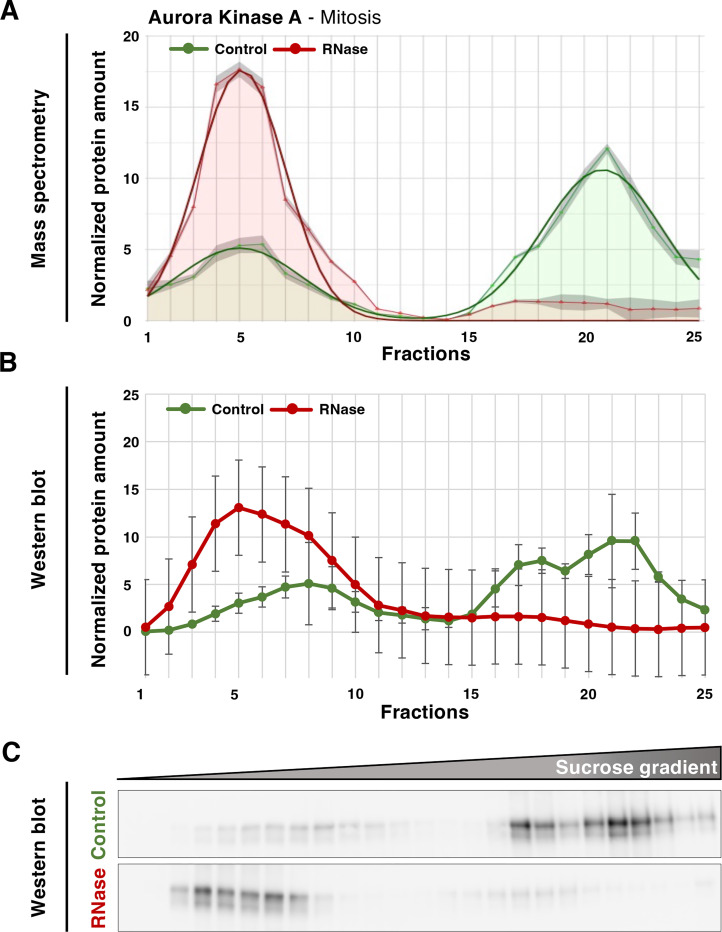
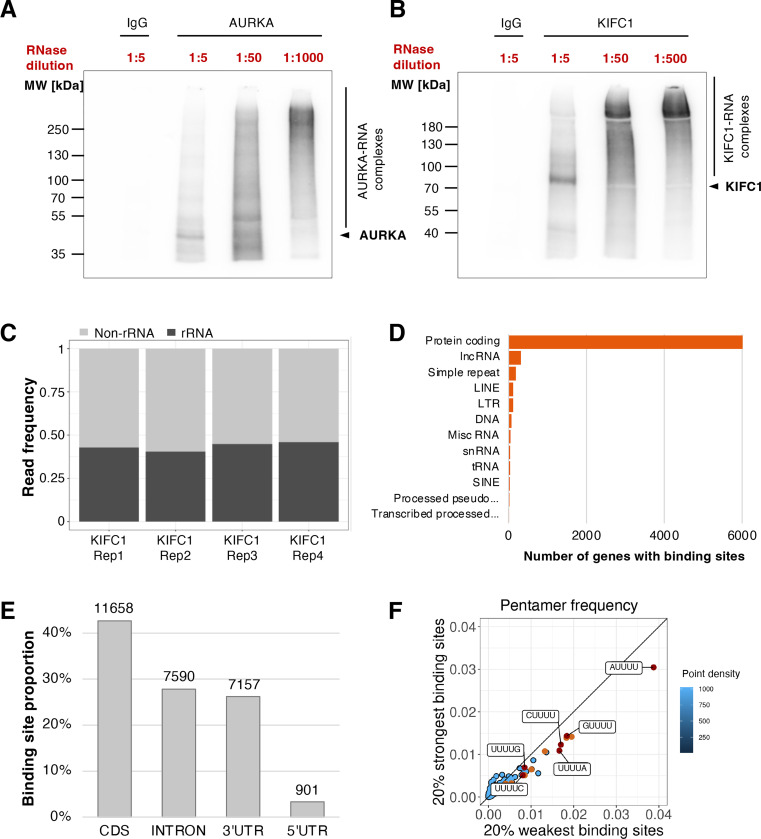
Ribonucleoprotein complexes are dynamic assemblies of RNA with RNA-binding proteins (RBPs), which can modulate the fate of the RNA molecules from transcription to degradation. Vice versa, RNA can regulate the interactions and functions of the associated proteins. Dysregulation of RBPs is linked to diseases such as cancer and neurological disorders. RNA and RBPs are present in mitotic structures like the centrosomes and spindle microtubules, but their influence on mitotic spindle integrity remains unknown. Thus, we applied the R-DeeP strategy for the proteome-wide identification of RNA-dependent proteins and complexes to cells synchronized in mitosis versus interphase. The resulting atlas of RNA-dependent proteins in cell division can be accessed through the R-DeeP 3.0 database (R-DeeP3.dkfz.de). It revealed key mitotic factors as RNA-dependent such as AURKA, KIFC1 and TPX2 that were linked to RNA despite their lack of canonical RNA-binding domains. KIFC1 was identified as a new interaction partner and phosphorylation substrate of AURKA at S349 and T359. In addition, KIFC1 interacted with both, AURKA and TPX2, in an RNA-dependent manner. Our data suggest a riboregulation of mitotic protein-protein interactions during spindle assembly, offering new perspectives on the control of cell division processes by RNA-protein complexes.
Keywords: RNA-binding proteins; mitotic factors; protein-protein interactions; riboregulation.
Conflict of interest statement
Declaration of interests S.D. is co-owner of siTOOLs Biotech, Martinsried, Germany, without relation to this work. The other authors disclose no conflicts of interest. This study is part of the PhD thesis of V.R.
Figures



References
-
- Dreyfuss G., Kim V.N. and Kataoka N. (2002) Messenger-RNA-binding proteins and the messages they carry. Nat Rev Mol Cell Biol, 3, 195–205. - PubMed
-
- Mitchell S.F. and Parker R. (2014) Principles and properties of eukaryotic mRNPs. Mol Cell, 54, 547–558. - PubMed
-
- Gebauer F., Schwarzl T., Valcarcel J. and Hentze M.W. (2021) RNA-binding proteins in human genetic disease. Nat Rev Genet, 22, 185–198. - PubMed
-
- Mayr C. (2017) Regulation by 3’-Untranslated Regions. Annu Rev Genet, 51, 171–194. - PubMed
-
- Hentze M.W., Castello A., Schwarzl T. and Preiss T. (2018) A brave new world of RNA-binding proteins. Nat Rev Mol Cell Biol, 19, 327–341. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous