

TRIM21 induces selective autophagic degradation of c-Myc and sensitizes regorafenib therapy in colorectal cancer
- PMID: 39388269
- PMCID: PMC11494295
- DOI: 10.1073/pnas.2406936121
TRIM21 induces selective autophagic degradation of c-Myc and sensitizes regorafenib therapy in colorectal cancer
Abstract
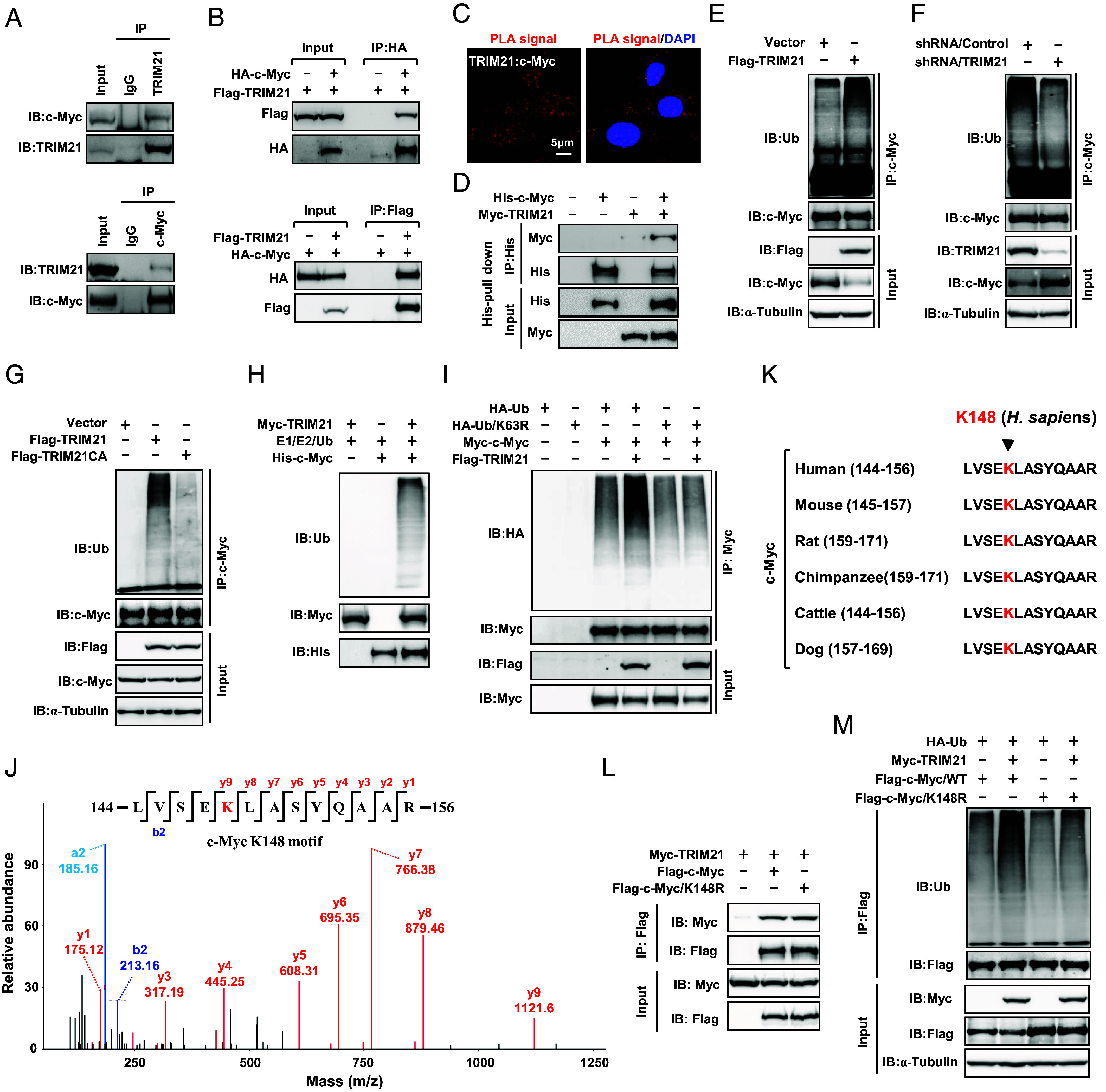
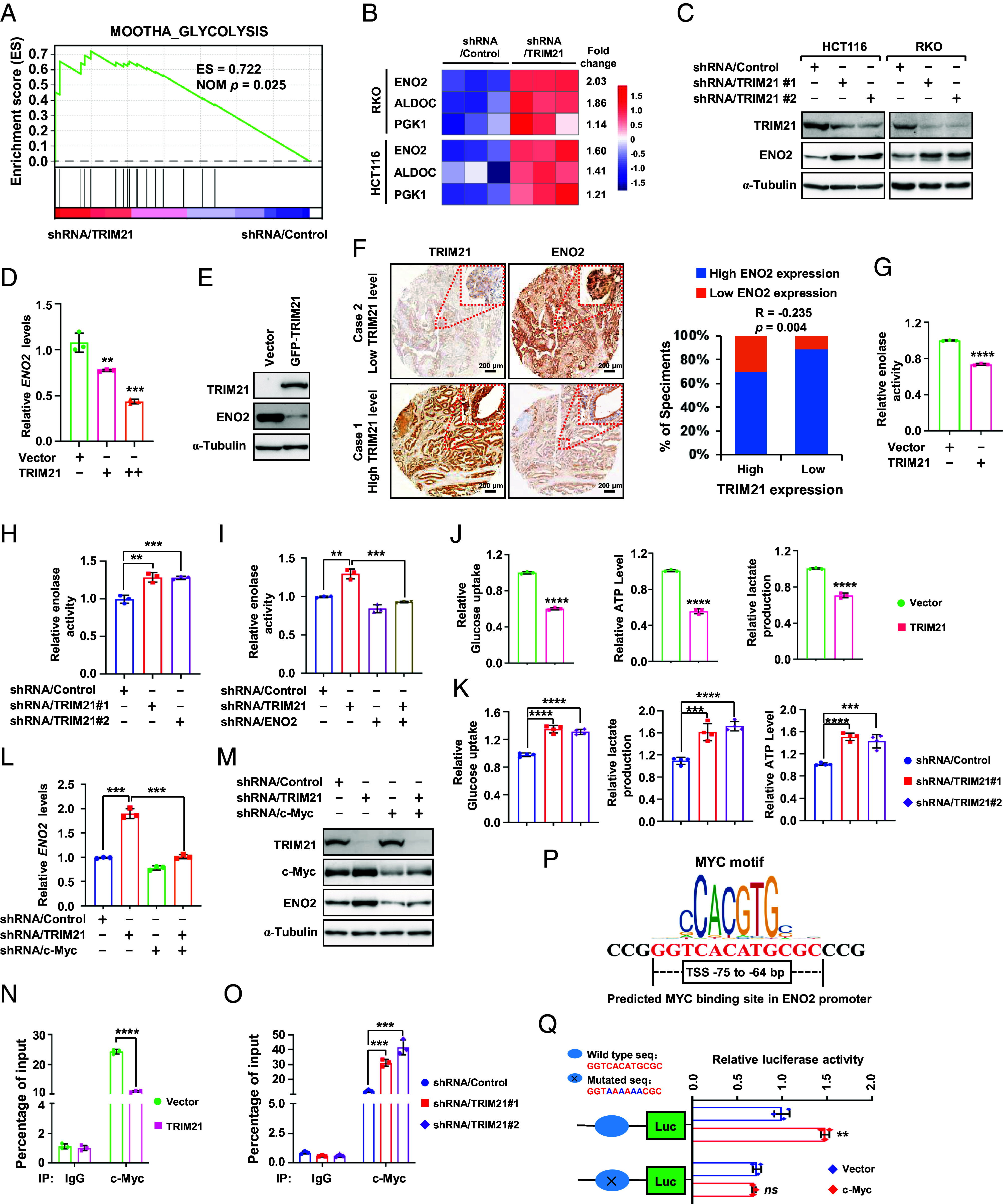
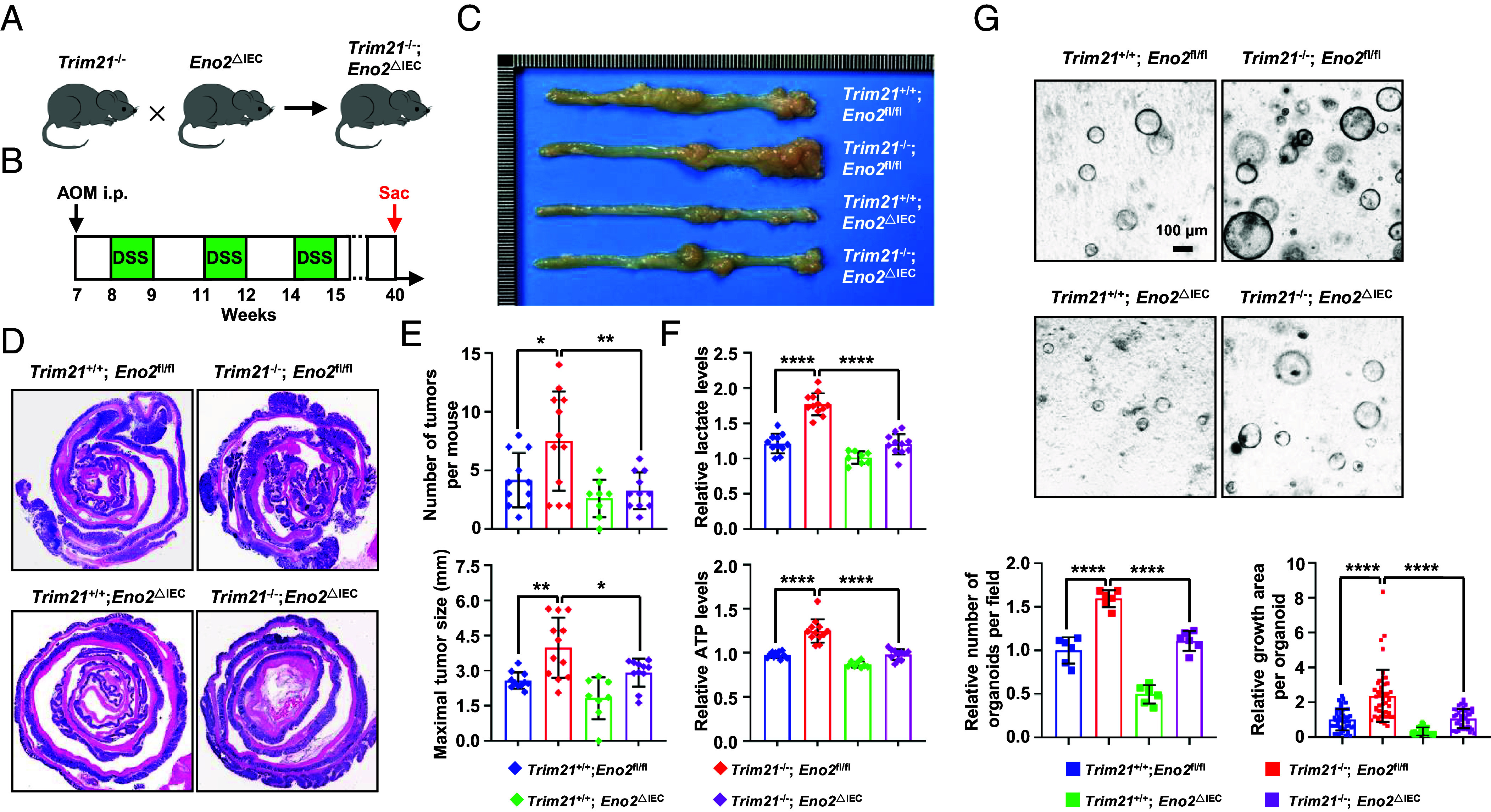
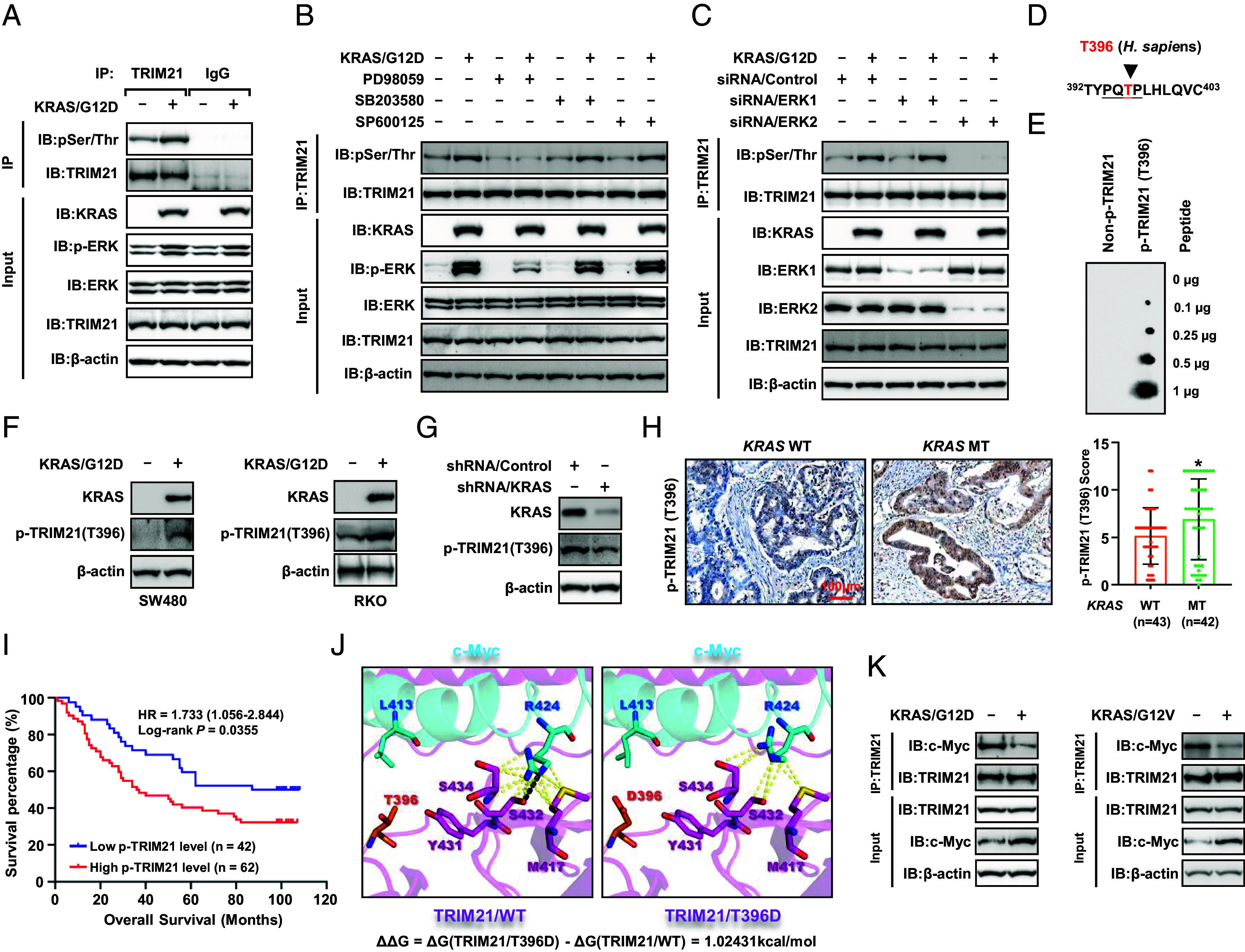
Kirsten rat sarcoma virus (KRAS) mutation is associated with malignant tumor transformation and drug resistance. However, the development of clinically effective targeted therapies for KRAS-mutant cancer has proven to be a formidable challenge. Here, we report that tripartite motif-containing protein 21 (TRIM21) functions as a target of extracellular signal-regulated kinase 2 (ERK2) in KRAS-mutant colorectal cancer (CRC), contributing to regorafenib therapy resistance. Mechanistically, TRIM21 directly interacts with and ubiquitinates v-myc avian myelocytomatosis viral oncogene homolog (c-Myc) at lysine 148 (K148) via K63-linkage, enabling c-Myc to be targeted to the autophagy machinery for degradation, ultimately resulting in the downregulation of enolase 2 expression and inhibition of glycolysis. However, mutant KRAS (KRAS/MT)-driven mitogen-activated protein kinase (MAPK) signaling leads to the phosphorylation of TRIM21 (p-TRIM21) at Threonine 396 (T396) by ERK2, disrupting the interaction between TRIM21 and c-Myc and thereby preventing c-Myc from targeting autophagy for degradation. This enhances glycolysis and contributes to regorafenib resistance. Clinically, high p-TRIM21 (T396) is associated with an unfavorable prognosis. Targeting TRIM21 to disrupt KRAS/MT-driven phosphorylation using the antidepressant vilazodone shows potential for enhancing the efficacy of regorafenib in treating KRAS-mutant CRC in preclinical models. These findings are instrumental for KRAS-mutant CRC treatment aiming at activating TRIM21-mediated selective autophagic degradation of c-Myc.
Keywords: KRAS; MYC; TRIM21; drug resistance; regorafenib.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures



References
MeSH terms
Substances
Grants and funding
- 82022050 82372662 81972601 82002520/National Natural Science Foundation of China
- 82373142/National Natural Science Foundation of China
- BE2023703/JST | Natural Science Foundation of Jiangsu Province (Jiangsu Natural Science Foundation)
- SZFCXK20241/Suzhou Medical Key Supported Discipline
- 82302967/National Natural Science Foundation of China
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
