

Optimized intravital three-photon imaging of intact mouse tibia links plasma cell motility to functional states
- PMID: 39391739
- PMCID: PMC11466647
- DOI: 10.1016/j.isci.2024.110985
Optimized intravital three-photon imaging of intact mouse tibia links plasma cell motility to functional states
Abstract
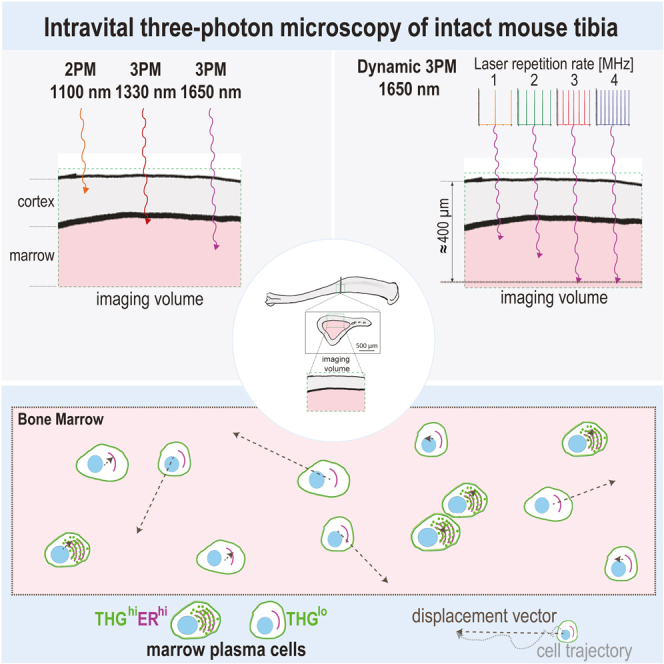
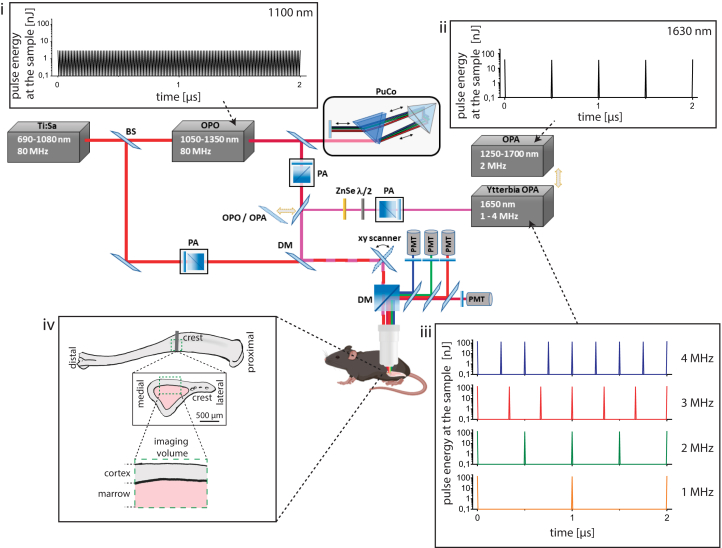
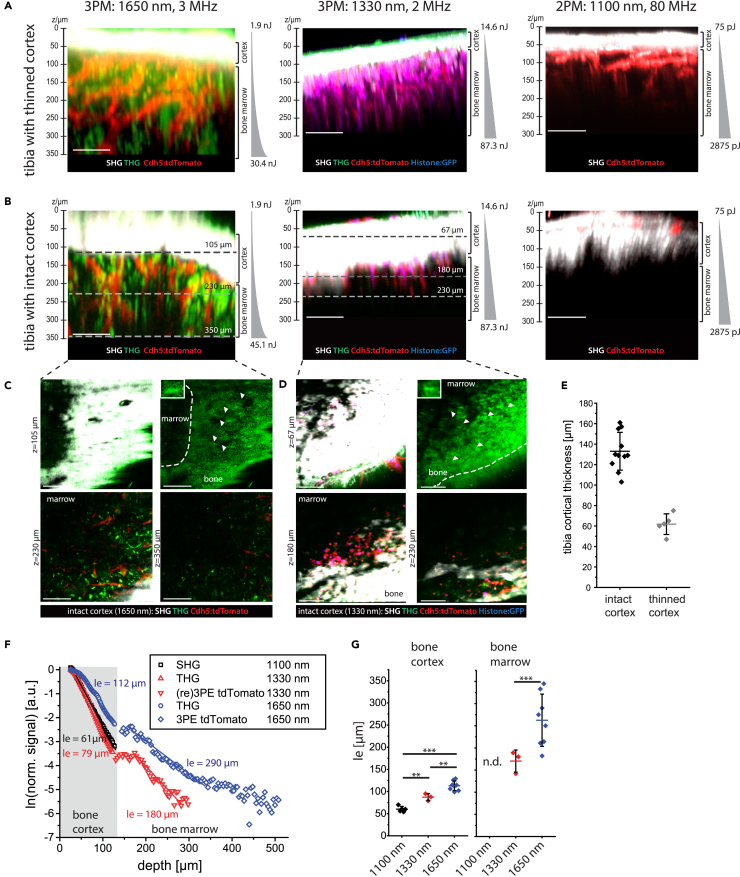
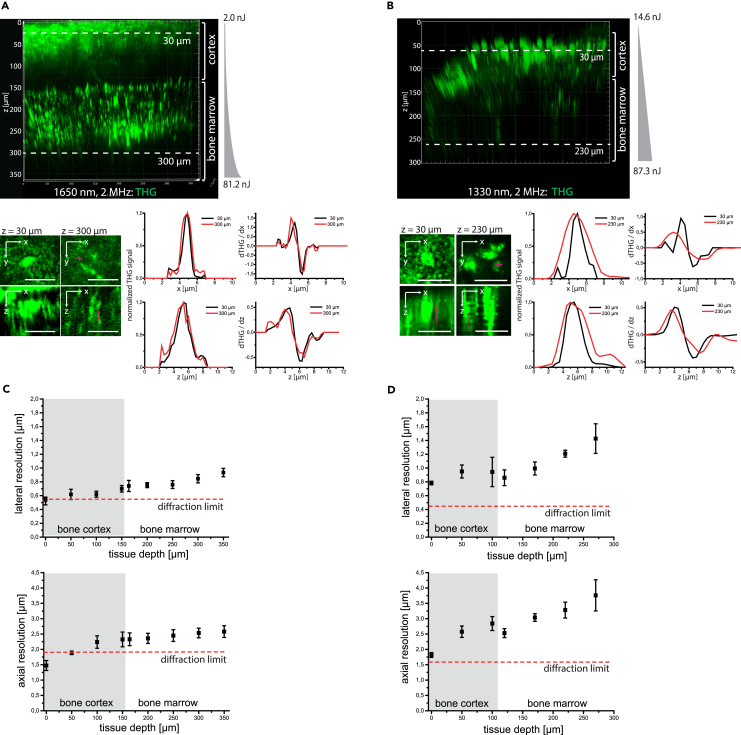
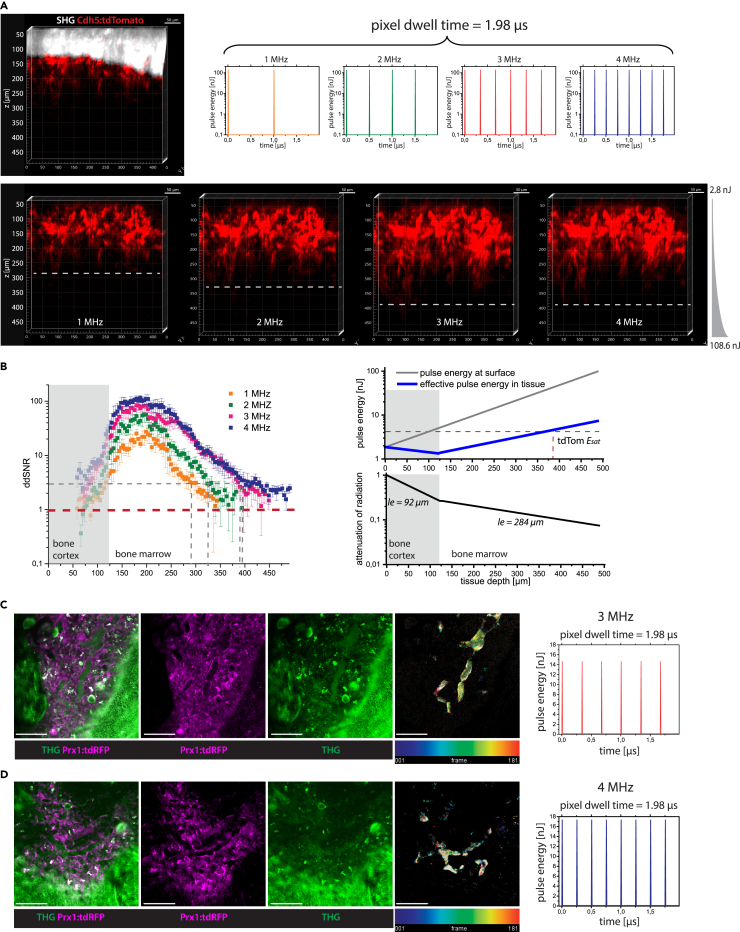
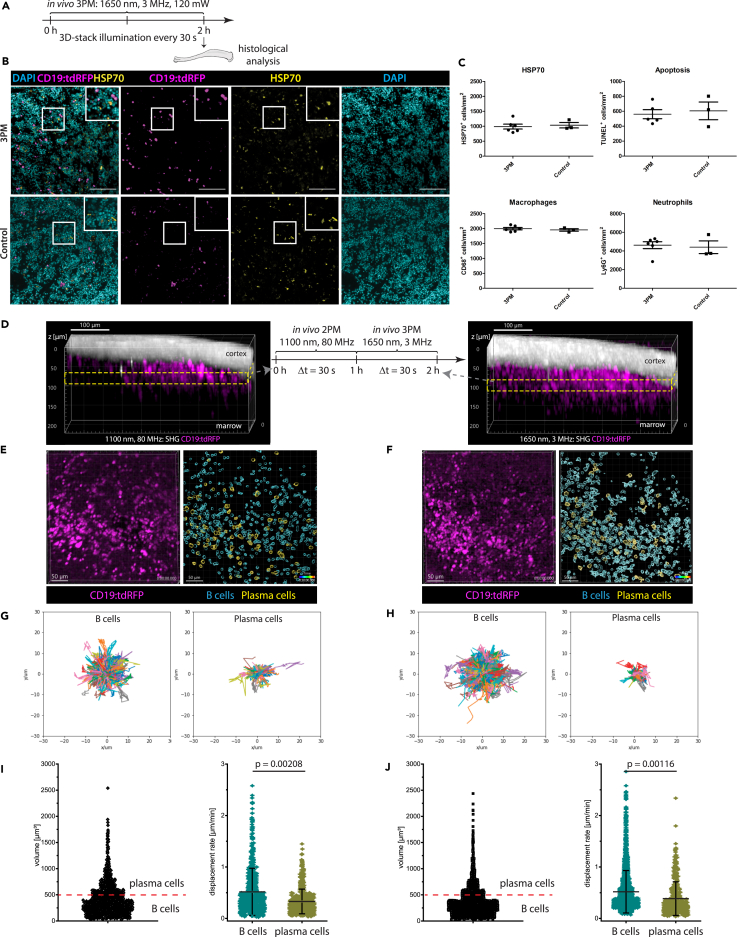
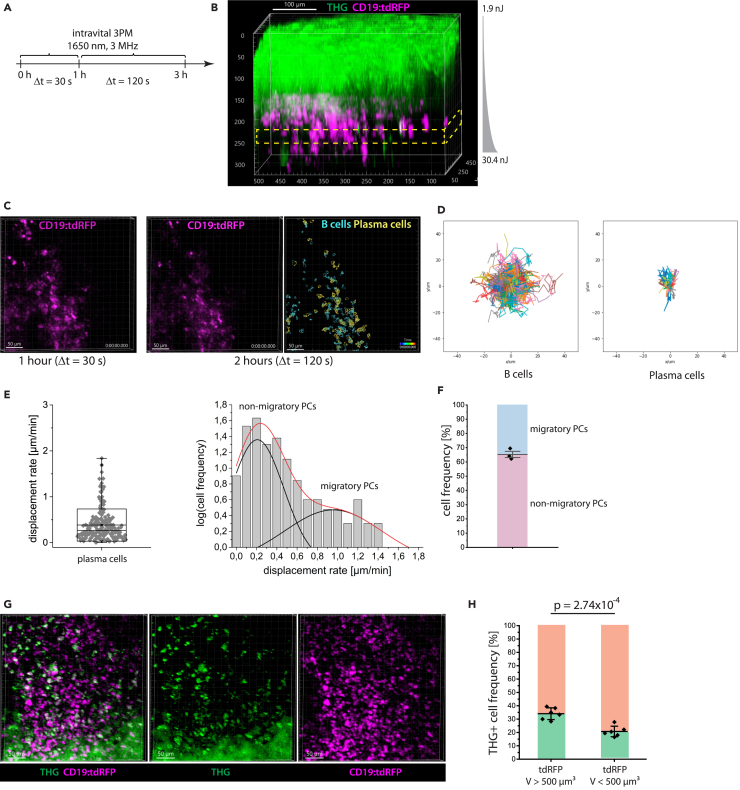
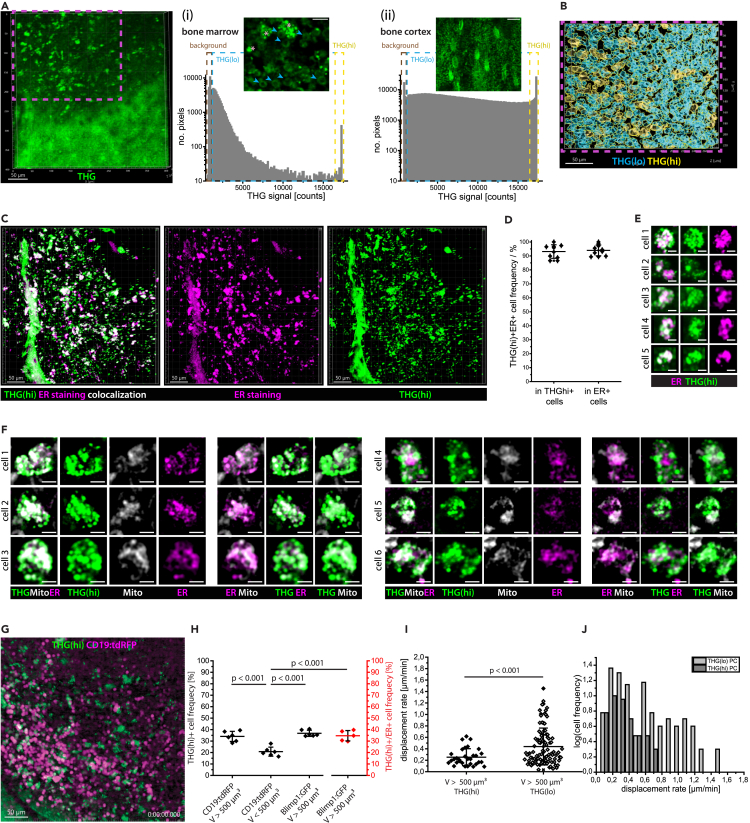
Intravital deep bone marrow imaging is crucial to studying cellular dynamics and functions but remains challenging, and minimally invasive methods are needed. We employed a high pulse-energy 1650 nm laser to perform three-photon microscopy in vivo, reaching ≈400 μm depth in intact mouse tibia. Repetition rates of 3 and 4 MHz allowed us to analyze motility patterns of fast and rare cells within unperturbed marrow and to identify a bi-modal migratory behavior for plasma cells. Third harmonic generation (THG) was identified as a label-free marker for cellular organelles, particularly endoplasmic reticulum, indicating protein synthesis capacity. We found a strong THG signal, suggesting high antibody secretion, in one-third of plasma cells while the rest showed low signals. We discovered an inverse relationship between migratory behavior and THG signal, linking motility to functional plasma cell states. This method may enhance our understanding of marrow microenvironment effects on cellular functions.
Keywords: Cell biology; Optical imaging; Small animal imaging.
© 2024 The Author(s).
Conflict of interest statement
S. Domingue and L. Wooldridge are with Thorlabs LASER Division CO, US. J. Heidelin and V. Andresen are with Miltenyi Biotec GmbH, Bergisch Gladbach, Germany.
Figures
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
