

Post-stroke hippocampal neurogenesis is impaired by microvascular dysfunction and PI3K signaling in cerebral amyloid angiopathy
- PMID: 39392753
- PMCID: PMC11562893
- DOI: 10.1016/j.celrep.2024.114848
Post-stroke hippocampal neurogenesis is impaired by microvascular dysfunction and PI3K signaling in cerebral amyloid angiopathy
Abstract
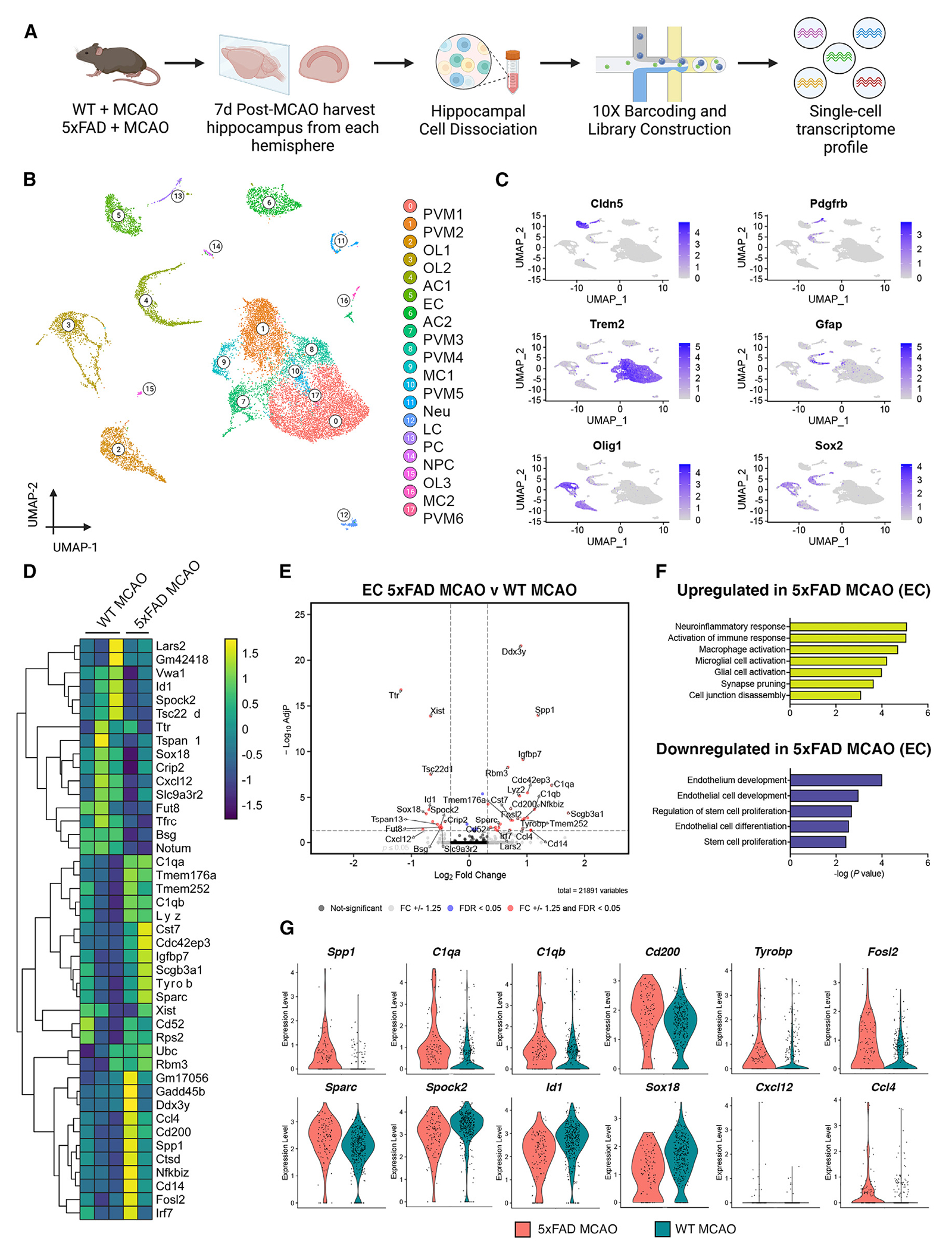
Ischemic stroke and cerebral amyloid angiopathy (CAA) pose significant challenges in an aging population, particularly in post-stroke recovery. Using the 5xFAD mouse model, we explore the relationship between CAA, ischemic stroke, and tissue recovery. We hypothesize that amyloid-beta accumulation worsens stroke outcomes by inducing blood-brain barrier (BBB) dysfunction, leading to impaired neurogenesis. Our findings show that CAA exacerbates stroke outcomes, with mice exhibiting constricted BBB microvessels, reduced cerebral blood flow, and impaired tissue recovery. Transcriptional analysis shows that endothelial cells and neural progenitor cells (NPCs) in the hippocampus exhibit differential gene expression in response to CAA and stroke, specifically targeting the phosphatidylinositol 3-kinase (PI3K) pathway. In vitro experiments with human NPCs validate these findings, showing that disruption of the CXCL12-PIK3C2A-CREB3L2 axis impairs neurogenesis. Notably, PI3K pathway activation restores neurogenesis, highlighting a potential therapeutic approach. These results suggest that CAA combined with stroke induces microvascular dysfunction and aberrant neurogenesis through this specific pathway.
Keywords: CAA; CP: Cell biology; CP: Neuroscience; PI3K; hippocampus; neurogenesis; stroke.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Similar articles
-
Endothelial Mitochondrial Dysfunction in Cerebral Amyloid Angiopathy and Alzheimer's Disease.J Alzheimers Dis. 2019;72(4):1019-1039. doi: 10.3233/JAD-190357. J Alzheimers Dis. 2019. PMID: 31306129 Free PMC article. Review.
-
Amyloid-β contributes to blood-brain barrier leakage in transgenic human amyloid precursor protein mice and in humans with cerebral amyloid angiopathy.Stroke. 2012 Feb;43(2):514-23. doi: 10.1161/STROKEAHA.111.627562. Epub 2011 Nov 23. Stroke. 2012. PMID: 22116809 Free PMC article.
-
Microvascular endothelial cells-derived microvesicles imply in ischemic stroke by modulating astrocyte and blood brain barrier function and cerebral blood flow.Mol Brain. 2016 Jun 7;9(1):63. doi: 10.1186/s13041-016-0243-1. Mol Brain. 2016. PMID: 27267759 Free PMC article.
-
Reduced nitric oxide bioavailability mediates cerebroarterial dysfunction independent of cerebral amyloid angiopathy in a mouse model of Alzheimer's disease.Am J Physiol Heart Circ Physiol. 2017 Feb 1;312(2):H232-H238. doi: 10.1152/ajpheart.00607.2016. Epub 2016 Nov 11. Am J Physiol Heart Circ Physiol. 2017. PMID: 27836896
-
From Stroke to Dementia: a Comprehensive Review Exposing Tight Interactions Between Stroke and Amyloid-β Formation.Transl Stroke Res. 2020 Aug;11(4):601-614. doi: 10.1007/s12975-019-00755-2. Epub 2019 Nov 28. Transl Stroke Res. 2020. PMID: 31776837 Free PMC article. Review.
Cited by
-
Models and mechanisms of post-stroke dementia and cognitive impairment.Front Stroke. 2025;4:1563924. doi: 10.3389/fstro.2025.1563924. Epub 2025 May 8. Front Stroke. 2025. PMID: 40757092 Free PMC article.
-
Vortioxetine alleviates motor, cognitive and emotional disorders in post-stroke rats by regulating the TLR-2/NF-κB pathway.Front Pharmacol. 2025 Mar 12;16:1555079. doi: 10.3389/fphar.2025.1555079. eCollection 2025. Front Pharmacol. 2025. PMID: 40144659 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
