

Neural activity for complex sounds in the marmoset anterior cingulate cortex
- PMID: 39394433
- PMCID: PMC11470068
- DOI: 10.1038/s42003-024-07019-2
Neural activity for complex sounds in the marmoset anterior cingulate cortex
Abstract
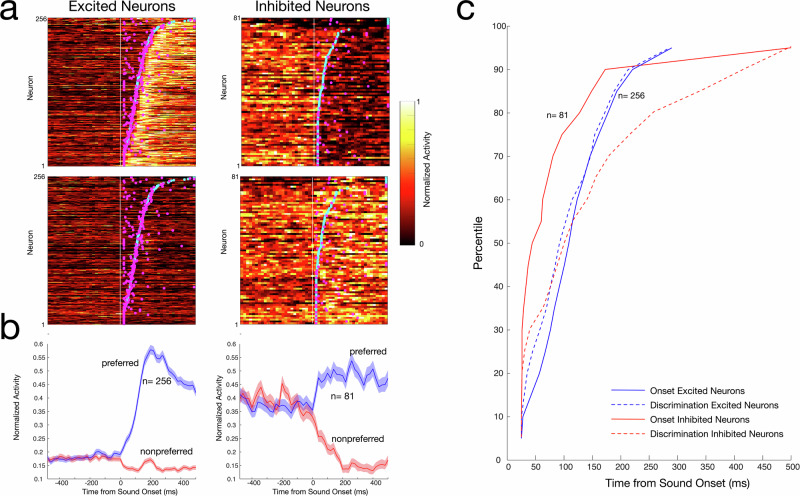
Vocalizations play an important role in the daily life of nonhuman primates and are likely precursors of human language. Recent functional imaging studies in the highly vocal common marmoset (Callithrix jacchus) have suggested that anterior cingulate cortex (ACC) area 32 may be a part of a vocalization-processing network but the response properties of area 32 neurons to auditory stimuli remain unknown. Here we perform electrophysiological recordings in area 32 in marmosets with high-density Neuropixels probes and characterize neuronal responses to a variety of sounds including conspecific vocalizations. Nearly half of the neurons in area 32 respond to conspecific vocalizations and other complex auditory stimuli. These responses exhibit dynamics consisting of an initially non-selective reduction in neural activity, followed by an increase in activity that immediately conveys sound selectivity. Our findings demonstrate that primate ACC area 32 processes species-specific and biologically relevant sounds.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Bezerra, B. M. & Souto, A. Structure and usage of the vocal repertoire of Callithrix jacchus. Int. J. Primatol.29, 671–701 (2008). - DOI
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
