

Arabidopsis MORC1 and MED9 Interact to Regulate Defense Gene Expression and Plant Fitness
- PMID: 39397299
- PMCID: PMC11471927
- DOI: 10.5423/PPJ.OA.07.2024.0107
Arabidopsis MORC1 and MED9 Interact to Regulate Defense Gene Expression and Plant Fitness
Abstract
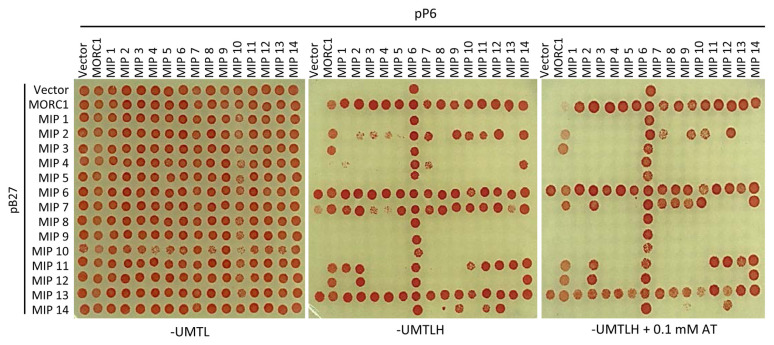
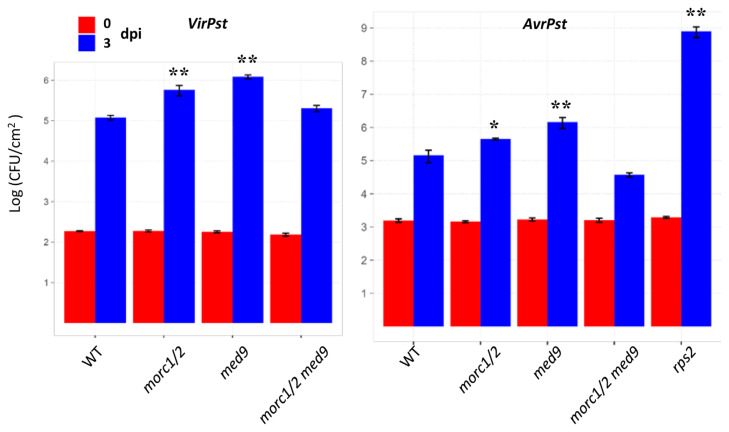
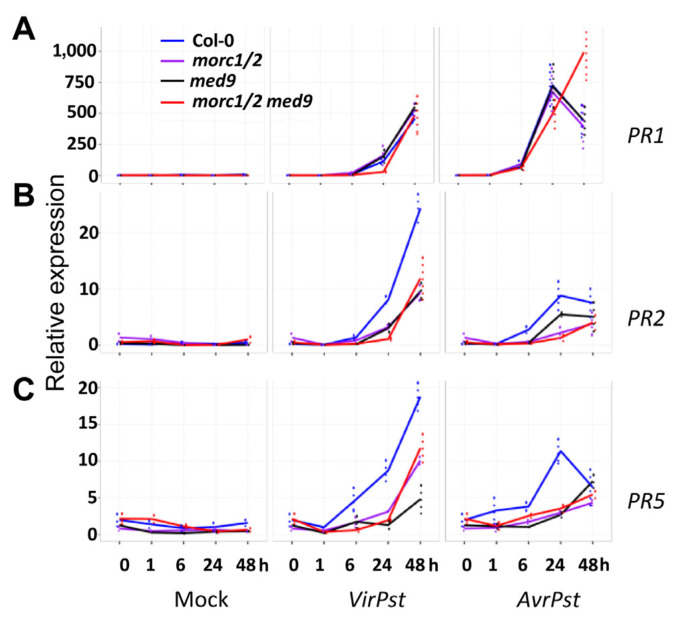
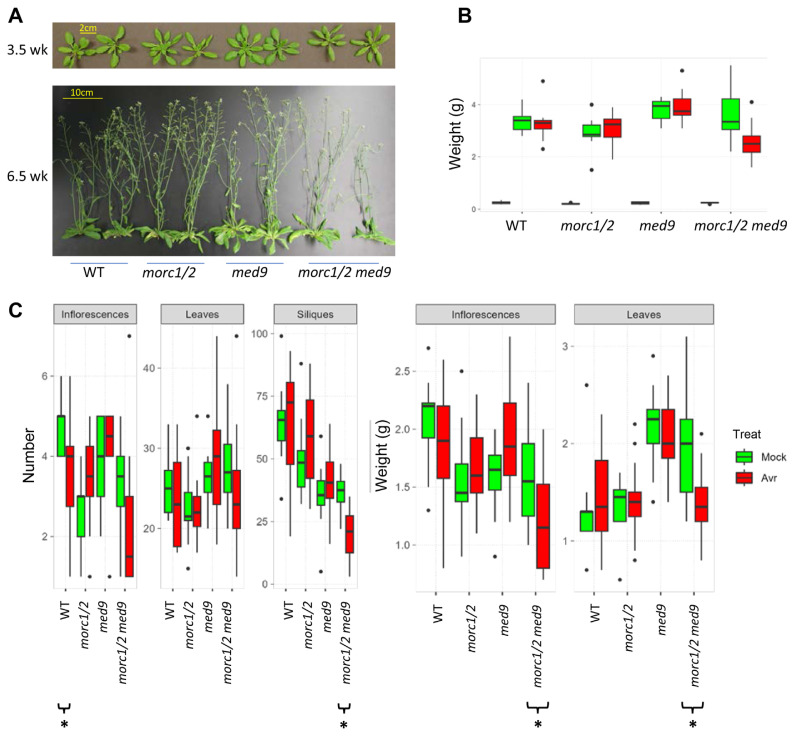
Arabidopsis MORC1 (Microrchidia) is required for multiple levels of immunity. We identified 14 MORC1-interacting proteins (MIPs) via yeast two-hybrid screening, eight of which have confirmed or putative nuclear-associated functions. While a few MIP mutants displayed altered bacterial resistance, MIP13 was unusual. The MIP13 mutant was susceptible to Pseudomonas syringae, but when combined with morc1/2, it regained wild-type resistance; notably, morc1/2 is susceptible to the same pathogen. MIP13 encodes MED9, a mediator complex component that interfaces with RNA polymerase II and transcription factors. Expression analysis of defense genes PR1, PR2, and PR5 in response to avirulent P. syringae revealed that morc1/2 med9 expressed these genes in a slow but sustained manner, unlike its lower-order mutants. This expression pattern may explain the restored resistance and suggests that the interplay of MORC1/2 and MED9 might be important in curbing defense responses to maintain fitness. Indeed, repeated challenges with avirulent P. syringae triggered significant growth inhibition in morc1/2 med9, indicating that MED9 and MORC1 may play an important role in balancing defense and growth. Furthermore, the in planta interaction of MED9 and MORC1 occurred 24 h, not 6 h, postinfection, suggesting that the interaction functions late in the defense signaling. Our study reveals a complex interplay between MORC1 and MED9 in maintaining an optimal balance between defense and growth in Arabidopsis.
Keywords: Arabidopsis; MORC1; defense; growth; mediator.
Conflict of interest statement
No potential conflict of interest relevant to this article was reported.
Figures



References
-
- Bassil E., Tajima H., Liang Y.-C., Ohto M.-A., Ushijima K., Nakano R., Esumi T., Coku A., Belmonte M., Blumwald E. The Arabidopsis Na+/H+ antiporters NHX1 and NHX2 control vacuolar pH and K+ homeostasis to regulate growth, flower development, and reproduction. Plant Cell. 2011;23:3482–3497. - PMC - PubMed
-
- Bonawitz N. D., Kim J. I., Tobimatsu Y., Ciesielski P. N., Anderson N. A., Ximenes E., Maeda J., Ralph J., Donohoe B. S., Ladisch M., Chapple C. Disruption of Mediator rescues the stunted growth of a lignin-deficient Arabidopsis mutant. Nature. 2014;509:376–380. - PubMed
-
- Bonfig K. B., Schreiber U., Gabler A., Roitsch T., Berger S. Infection with virulent and avirulent P. syringae strains differentially affects photosynthesis and sink metabolism in Arabidopsis leaves. Planta. 2006;225:1–12. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
