

Cytokinetic contractile ring structural progression in an early embryo: positioning of scaffolding proteins, recruitment of α-actinin, and effects of myosin II inhibition
- PMID: 39398481
- PMCID: PMC11467475
- DOI: 10.3389/fcell.2024.1483345
Cytokinetic contractile ring structural progression in an early embryo: positioning of scaffolding proteins, recruitment of α-actinin, and effects of myosin II inhibition
Abstract
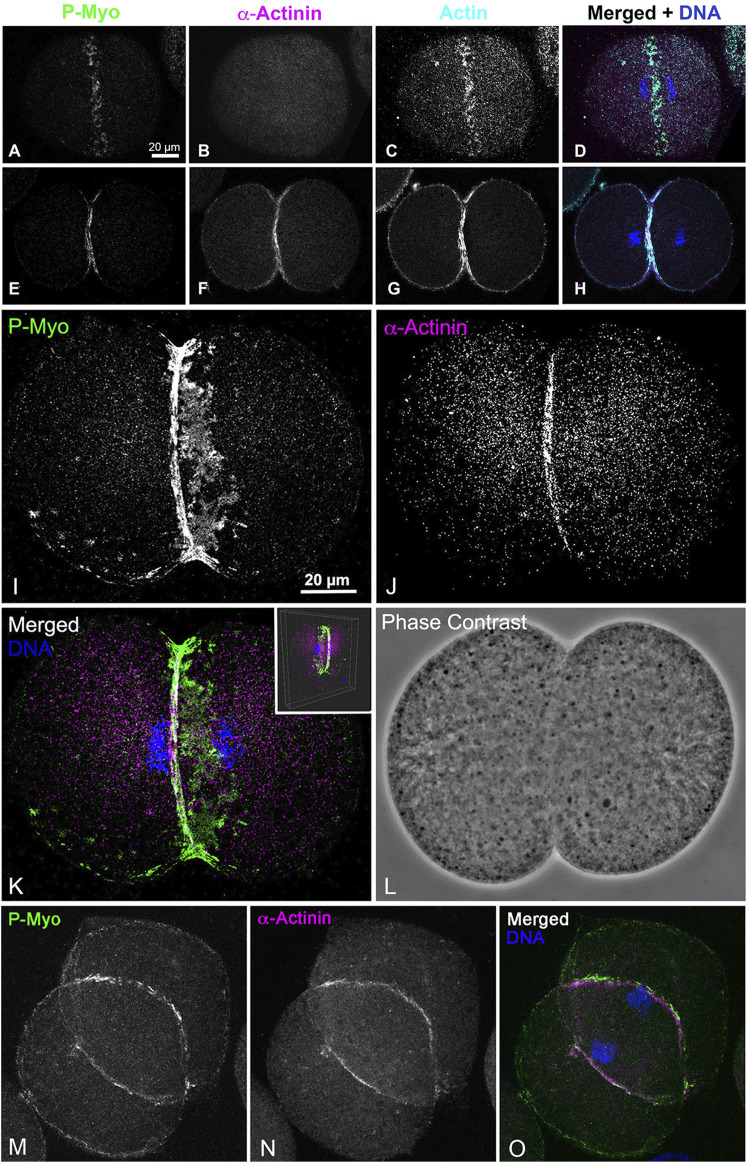
Our knowledge of the assembly and dynamics of the cytokinetic contractile ring (CR) in animal cells remains incomplete. We have previously used super-resolution light microscopy and platinum replica electron microscopy to elucidate the ultrastructural organization of the CR in first division sea urchin embryos. To date, our studies indicate that the CR initiates as an equatorial band of clusters containing myosin II, actin, septin and anillin, which then congress over time into patches which coalesce into a linear array characteristic of mature CRs. In the present study, we applied super-resolution interferometric photoactivated localization microscopy to confirm the existence of septin filament-like structures in the developing CR, demonstrate the close associations between septin2, anillin, and myosin II in the CR, as well as to show that septin2 appears consistently submembranous, whereas anillin is more widely distributed in the early CR. We also provide evidence that the major actin cross-linking protein α-actinin only associates with the linearized, late-stage CR and not with the early CR clusters, providing further support to the idea that α-actinin associates with actomyosin structures under tension and can serve as a counterbalance. In addition, we show that inhibition of actomyosin contraction does not stop the assembly of the early CR clusters but does arrest the progression of these structures to the aligned arrays required for functional cytokinesis. Taken together our results reinforce and extend our model for a cluster to patch to linear structural progression of the CR in sea urchin embryos and highlight the evolutionary relationships with cytokinesis in fission yeast.
Keywords: actin; anillin; contractile ring; cytokinesis; myosin II; sea urchin; septin; α-actinin.
Copyright © 2024 Henson, Reyes, Lo, Herrera, McKim, Herzon, Galvez-Ceron, Hershey, Kim and Shuster.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Figures








References
LinkOut - more resources
Full Text Sources