

Dietary ellagic acid blocks inflammation-associated atherosclerotic plaque formation in cholesterol-fed apoE-deficient mice
- PMID: 39398881
- PMCID: PMC11464280
- DOI: 10.4162/nrp.2024.18.5.617
Dietary ellagic acid blocks inflammation-associated atherosclerotic plaque formation in cholesterol-fed apoE-deficient mice
Abstract
Background/objectives: Atherosclerosis particularly due to high circulating level of low-density lipoprotein is a major cause of cardiovascular diseases. Ellagic acid is a natural polyphenolic compound rich in pomegranates and berries. Our previous study showed that ellagic acid improved functionality of reverse cholesterol transport in murine model of atherosclerosis. The aim of this study is to investigate whether ellagic acid inhibited inflammation-associated atherosclerotic plaque formation in cholesterol-fed apolipoprotein E (apoE)-knockout (KO) mice.
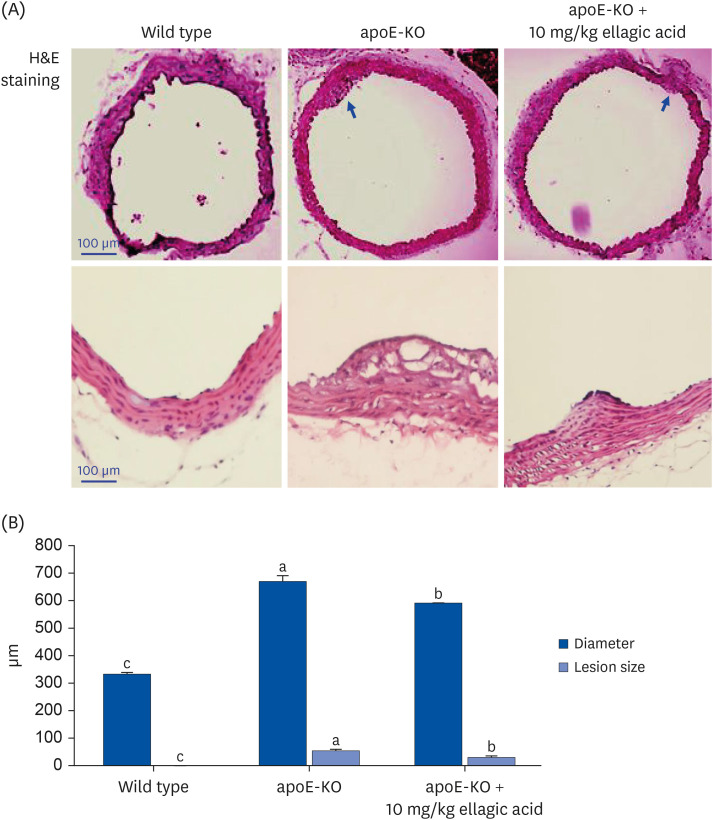
Materials/methods: Wild type mice and apoE-KO mice were fed a cholesterol-rich Paigen diet for 10 weeks to induce severe atherosclerosis. Concurrently, 10 mg/kg ellagic acid was orally administered to the apoE-KO mice. Plaque lesion formation and lipid deposition were examined by staining with hematoxylin and eosin, Sudan IV and oil red O.
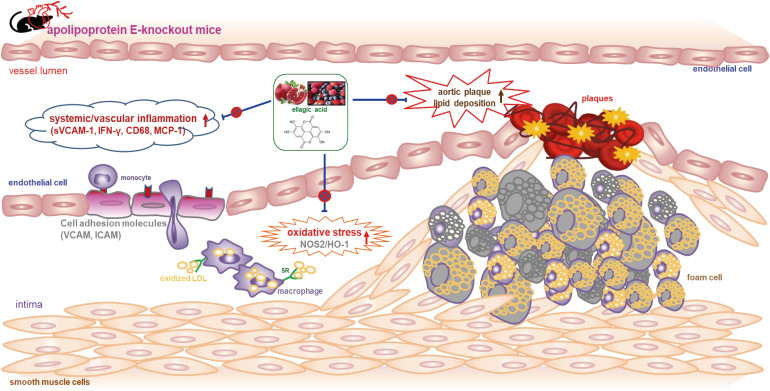
Results: The plasma leukocyte profile of cholesterol-fed mice was not altered by apoE deficiency. Oral administration of ellagic acid attenuated plaque lesion formation and lipid deposition in the aorta tree of apoE-KO mice. Ellagic acid substantially reduced plasma levels of soluble vascular cell adhesion molecule and interferon-γ in Paigen diet-fed apoE-KO mice. When 10 mg/kg ellagic acid was administered to cholesterol-fed apoE-KO mice, the levels of CD68 and MCP-1 were strongly reduced in aorta vessels. The protein expression level of nitric oxide synthase-2 (NOS2) in the aorta was highly enhanced by supplementation of ellagic acid to apoE-KO mice, but the expression level of heme oxygenase-1 (HO-1) in the aorta was reduced. Furthermore, ellagic acid diminished the increased aorta expression of the inflammatory adhesion molecules in cholesterol-fed apoE-KO mice. The treatment of ellagic acid inhibited the scavenger receptor-B1 expression in the aorta of apoE-KO mice, while the cholesterol efflux-related transporters were not significantly changed.
Conclusion: These results suggest that ellagic acid may be an atheroprotective compound by attenuating apoE deficiency-induced vascular inflammation and reducing atherosclerotic plaque lesion formation.
Keywords: Apolipoproteins E; atherosclerosis; atherosclerotic plaque; ellagic acid; inflammation.
©2024 The Korean Nutrition Society and the Korean Society of Community Nutrition.
Conflict of interest statement
Conflict of Interest: The authors declare that they have no potential conicts of interests.
Figures






References
-
- Fazio S, Linton MF. Mouse models of hyperlipidemia and atherosclerosis. Front Biosci. 2001;6:D515–D525. - PubMed
-
- Meir KS, Leitersdorf E. Atherosclerosis in the apolipoprotein-E-deficient mouse: a decade of progress. Arterioscler Thromb Vasc Biol. 2004;24:1006–1014. - PubMed
-
- Nakashima Y, Plump AS, Raines EW, Breslow JL, Ross R. ApoE-deficient mice develop lesions of all phases of atherosclerosis throughout the arterial tree. Arterioscler Thromb. 1994;14:133–140. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
