

Induction of long-term hyperexcitability by memory-related cAMP signaling in isolated nociceptor cell bodies
- PMID: 39399224
- PMCID: PMC11470187
- DOI: 10.1016/j.ynpai.2024.100166
Induction of long-term hyperexcitability by memory-related cAMP signaling in isolated nociceptor cell bodies
Abstract
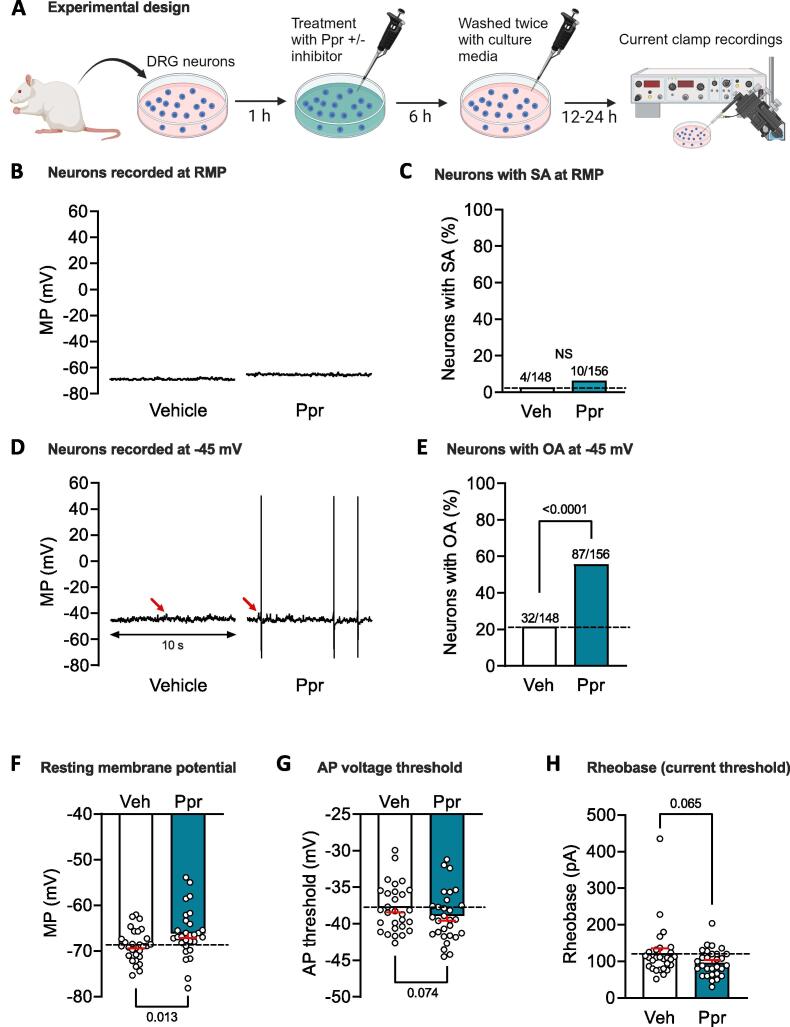
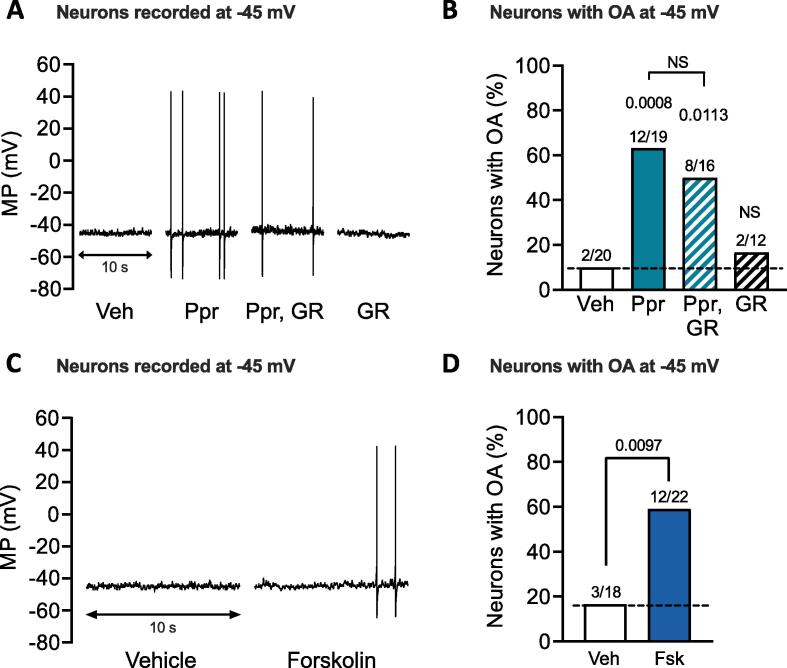
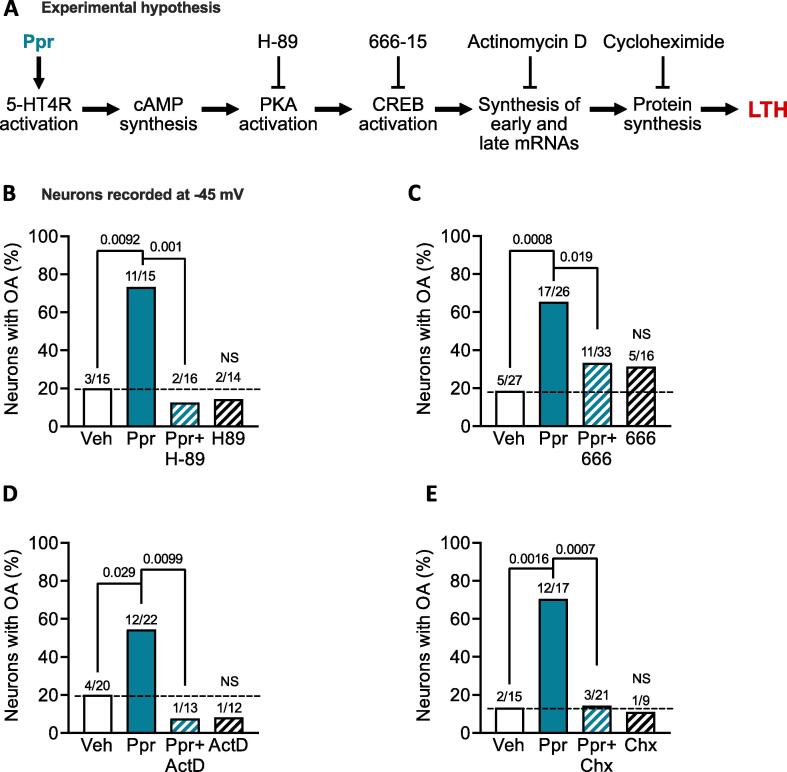
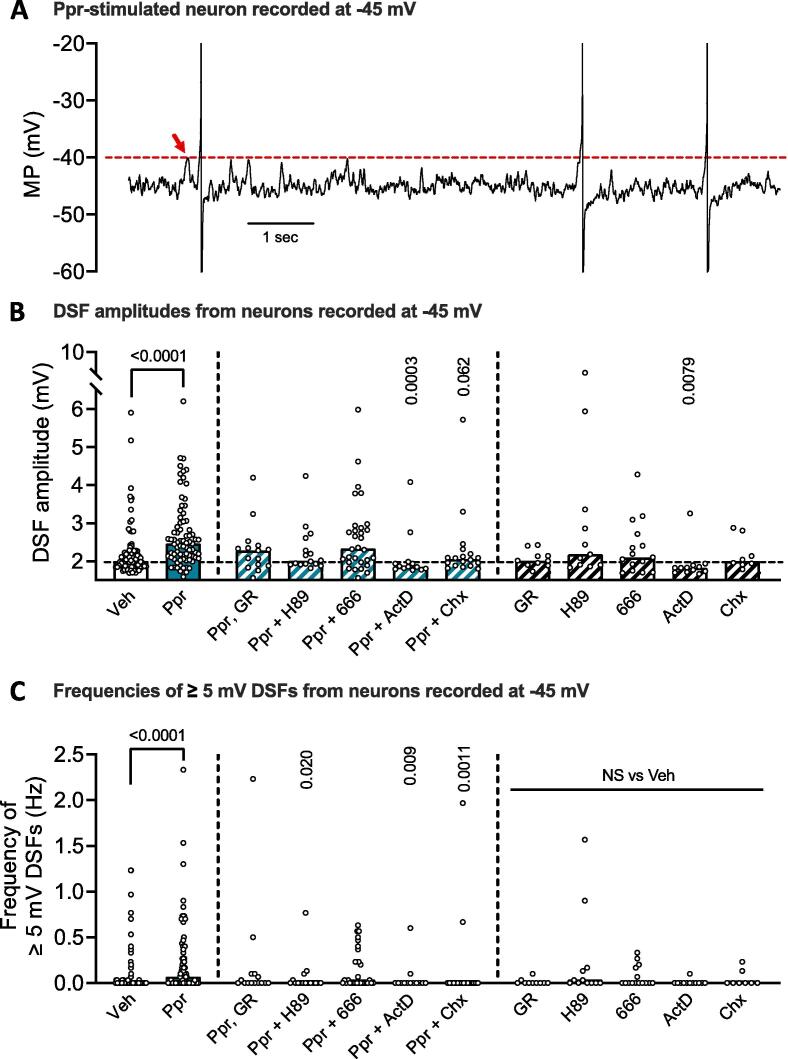
Persistent hyperactivity of nociceptors is known to contribute significantly to long-lasting sensitization and ongoing pain in many clinical conditions. It is often assumed that nociceptor hyperactivity is mainly driven by continuing stimulation from inflammatory mediators. We have tested an additional possibility: that persistent increases in excitability promoting hyperactivity can be induced by a prototypical cellular signaling pathway long known to induce late-phase long-term potentiation (LTP) of synapses in brain regions involved in memory formation. This cAMP-PKA-CREB-gene transcription-protein synthesis pathway was tested using whole-cell current clamp methods on small dissociated sensory neurons (primarily nociceptors) from dorsal root ganglia (DRGs) excised from previously uninjured ("naïve") male rats. Six-hour treatment with the specific Gαs-coupled 5-HT4 receptor agonist, prucalopride, or with the adenylyl cyclase activator forskolin induced long-term hyperexcitability (LTH) in DRG neurons that manifested 12-24 h later as action potential (AP) discharge (ongoing activity, OA) during artificial depolarization to -45 mV, a membrane potential that is normally subthreshold for AP generation. Prucalopride treatment also induced significant long-lasting depolarization of resting membrane potential (from -69 to -66 mV), enhanced depolarizing spontaneous fluctuations (DSFs) of membrane potential, and produced trends for reduced AP threshold and rheobase. LTH was prevented by co-treatment of prucalopride with inhibitors of PKA, CREB, gene transcription, or protein synthesis. As in the induction of synaptic memory, many other cellular signals are likely to be involved. However, the discovery that this prototypical memory induction pathway can induce nociceptor LTH, along with reports that cAMP signaling and CREB activity in DRGs can induce hyperalgesic priming, suggest that early, temporary, cAMP-induced transcriptional and translational mechanisms can induce nociceptor LTH that might last for long periods. The present results also raise the question of whether reactivation of primed signaling mechanisms by re-exposure to inflammatory mediators linked to cAMP synthesis during subsequent challenges to bodily integrity can "reconsolidate" nociceptor memory, extending the duration of persistent hyperexcitability.
Keywords: Cellular memory; Cyclic AMP signaling; Dorsal root ganglia; Excitability; Hyperexcitability; Inflammatory mediator.
© 2024 The Authors.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Update of
-
Induction of long-term hyperexcitability by memory-related cAMP signaling in isolated nociceptor cell bodies.bioRxiv [Preprint]. 2024 Jul 17:2024.07.13.603393. doi: 10.1101/2024.07.13.603393. bioRxiv. 2024. Update in: Neurobiol Pain. 2024 Sep 20;16:100166. doi: 10.1016/j.ynpai.2024.100166. PMID: 39071414 Free PMC article. Updated. Preprint.
References
-
- Abram S.E., Yi J., Fuchs A., Hogan Q.H. Permeability of injured and intact peripheral nerves and dorsal root ganglia. Anesthesiology. 2006;105:146–153. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
