

A role for cross-linking proteins in actin filament network organization and force generation
- PMID: 39405356
- PMCID: PMC11513903
- DOI: 10.1073/pnas.2407838121
A role for cross-linking proteins in actin filament network organization and force generation
Abstract
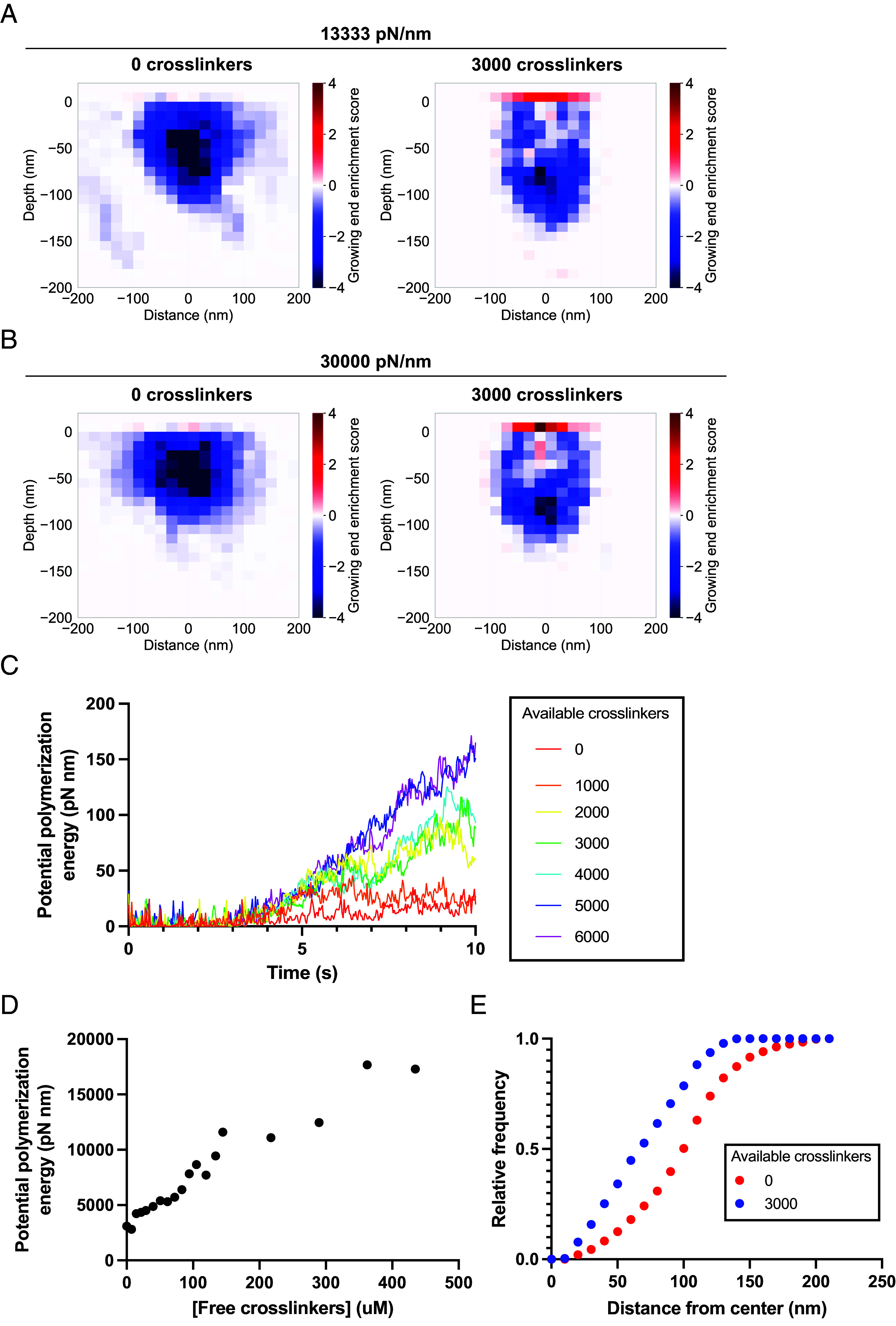
The high turgor pressure across the plasma membrane of yeasts creates a requirement for substantial force production by actin polymerization and myosin motor activity for clathrin-mediated endocytosis (CME). Endocytic internalization is severely impeded in the absence of fimbrin, an actin filament crosslinking protein called Sac6 in budding yeast. Here, we combine live-cell imaging and mathematical modeling to gain insights into the role of actin filament crosslinking proteins in force generation. Genetic manipulation showed that CME sites with more crosslinking proteins are more effective at internalization under high load. Simulations of an experimentally constrained, agent-based mathematical model recapitulate the result that endocytic networks with more double-bound fimbrin molecules internalize the plasma membrane against elevated turgor pressure more effectively. Networks with large numbers of crosslinks also have more growing actin filament barbed ends at the plasma membrane, where the addition of new actin monomers contributes to force generation and vesicle internalization. Our results provide a richer understanding of the crucial role played by actin filament crosslinking proteins during actin network force generation, highlighting the contribution of these proteins to the self-organization of the actin filament network and force generation under increased load.
Keywords: actin; clathrin-mediated endocytosis; crosslinking proteins; mathematical modeling.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures




Update of
-
A Role for Cross-linking Proteins in Actin Filament Network Organization and Force Generation.bioRxiv [Preprint]. 2024 Apr 20:2024.04.19.590161. doi: 10.1101/2024.04.19.590161. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2024 Oct 22;121(43):e2407838121. doi: 10.1073/pnas.2407838121. PMID: 38659919 Free PMC article. Updated. Preprint.
References
-
- Kaksonen M., Toret C. P., Drubin D. G., A modular design for the clathrin- and actin-mediated endocytosis machinery. Cell 123, 305–320 (2005). - PubMed
-
- Merrifield C. J., Feldman M. E., Wan L., Almers W., Imaging actin and dynamin recruitment during invagination of single clathrin-coated pits. Nat. Cell Biol. 4, 691–698 (2002). - PubMed
-
- Kaksonen M., Sun Y., Drubin D. G., A pathway for association of receptors, adaptors, and actin during endocytic internalization. Cell 115, 475–487 (2003). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
