

Sall4 and Gata4 induce cardiac fibroblast transition towards a partially multipotent state with cardiogenic potential
- PMID: 39406776
- PMCID: PMC11480346
- DOI: 10.1038/s41598-024-73975-8
Sall4 and Gata4 induce cardiac fibroblast transition towards a partially multipotent state with cardiogenic potential
Abstract
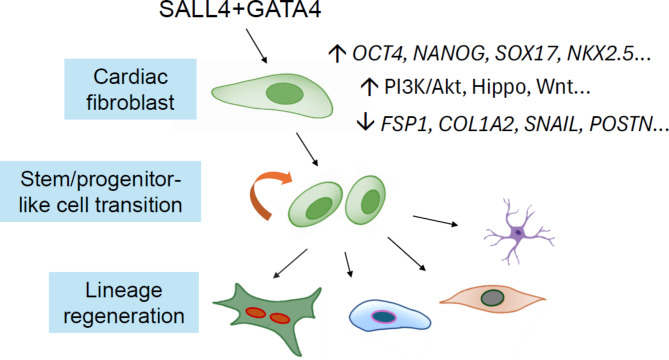
Cardiac cellular fate transition holds remarkable promise for the treatment of ischemic heart disease. We report that overexpressing two transcription factors, Sall4 and Gata4, which play distinct and overlapping roles in both pluripotent stem cell reprogramming and embryonic heart development, induces a fraction of stem-like cells in rodent cardiac fibroblasts that exhibit unlimited ex vivo expandability with clonogenicity. Transcriptomic and phenotypic analyses reveal that around 32 ± 6.4% of the expanding cells express Nkx2.5, while 13 ± 3.6% express Oct4. Activated signaling pathways like PI3K/Akt, Hippo, Wnt, and multiple epigenetic modification enzymes are also detected. Under suitable conditions, these cells demonstrate a high susceptibility to differentiating into cardiomyocyte, endothelial cell, and extracardiac neuron-like cells. The presence of partially pluripotent-like cells is characterized by alkaline phosphatase staining, germ layer marker expression, and tumor formation in injected mice (n = 5). Additionally, significant stem-like fate transitions and cardiogenic abilities are induced in human cardiac fibroblasts, but not in rat or human skin fibroblasts. Molecularly, we identify that SALL4 and GATA4 physically interact and synergistically stimulate the promoters of pluripotency genes but repress fibrogenic gene, which correlates with a primitive transition process. Together, this study uncovers a new cardiac regenerative mechanism that could potentially advance therapeutic endeavors and tissue engineering.
Keywords: Cardiac regeneration; Oct4; Partial reprogramming; Pluripotency; Protein interaction; Sox17.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures








References
-
- Cahill, T. J., Choudhury, R. P. & Riley, P. R. Heart regeneration and repair after myocardial infarction: translational opportunities for novel therapeutics. Nat. Rev. Drug Discov. 16, 699–717. 10.1038/nrd.2017.106 (2017). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
