

Unveiling lignocellulolytic potential: a genomic exploration of bacterial lineages within the termite gut
- PMID: 39407345
- PMCID: PMC11481507
- DOI: 10.1186/s40168-024-01917-7
Unveiling lignocellulolytic potential: a genomic exploration of bacterial lineages within the termite gut
Abstract
Background: The microbial landscape within termite guts varies across termite families. The gut microbiota of lower termites (LT) is dominated by cellulolytic flagellates that sequester wood particles in their digestive vacuoles, whereas in the flagellate-free higher termites (HT), cellulolytic activity has been attributed to fiber-associated bacteria. However, little is known about the role of individual lineages in fiber digestion, particularly in LT.
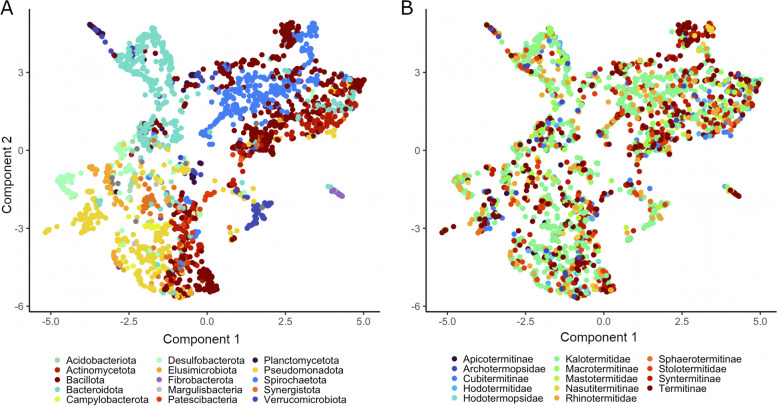
Results: We investigated the lignocellulolytic potential of 2223 metagenome-assembled genomes (MAGs) recovered from the gut metagenomes of 51 termite species. In the flagellate-dependent LT, cellulolytic enzymes are restricted to MAGs of Bacteroidota (Dysgonomonadaceae, Tannerellaceae, Bacteroidaceae, Azobacteroidaceae) and Spirochaetota (Breznakiellaceae) and reflect a specialization on cellodextrins, whereas their hemicellulolytic arsenal features activities on xylans and diverse heteropolymers. By contrast, the MAGs derived from flagellate-free HT possess a comprehensive arsenal of exo- and endoglucanases that resembles that of termite gut flagellates, underlining that Fibrobacterota and Spirochaetota occupy the cellulolytic niche that became vacant after the loss of the flagellates. Furthermore, we detected directly or indirectly oxygen-dependent enzymes that oxidize cellulose or modify lignin in MAGs of Pseudomonadota (Burkholderiales, Pseudomonadales) and Actinomycetota (Actinomycetales, Mycobacteriales), representing lineages located at the hindgut wall.
Conclusions: The results of this study refine our concept of symbiotic digestion of lignocellulose in termite guts, emphasizing the differential roles of specific bacterial lineages in both flagellate-dependent and flagellate-independent breakdown of cellulose and hemicelluloses, as well as a so far unappreciated role of oxygen in the depolymerization of plant fiber and lignin in the microoxic periphery during gut passage in HT. Video Abstract.
Keywords: CAZymes; Cellulase; Functional genomics; Lignin; Lignocellulose degradation; Termite microbiota.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






Similar articles
-
Comparison of microbial diversity and carbohydrate-active enzymes in the hindgut of two wood-feeding termites, Globitermes sulphureus (Blattaria: Termitidae) and Coptotermes formosanus (Blattaria: Rhinotermitidae).BMC Microbiol. 2024 Nov 12;24(1):470. doi: 10.1186/s12866-024-03623-8. BMC Microbiol. 2024. PMID: 39533168 Free PMC article.
-
Genome reduction and horizontal gene transfer in the evolution of Endomicrobia-rise and fall of an intracellular symbiosis with termite gut flagellates.mBio. 2024 Jun 12;15(6):e0082624. doi: 10.1128/mbio.00826-24. Epub 2024 May 14. mBio. 2024. PMID: 38742878 Free PMC article.
-
Fiber-associated spirochetes are major agents of hemicellulose degradation in the hindgut of wood-feeding higher termites.Proc Natl Acad Sci U S A. 2018 Dec 18;115(51):E11996-E12004. doi: 10.1073/pnas.1810550115. Epub 2018 Nov 30. Proc Natl Acad Sci U S A. 2018. PMID: 30504145 Free PMC article.
-
The Gut Microbiota of Termites: Digesting the Diversity in the Light of Ecology and Evolution.Annu Rev Microbiol. 2015;69:145-66. doi: 10.1146/annurev-micro-092412-155715. Epub 2015 Jul 16. Annu Rev Microbiol. 2015. PMID: 26195303 Review.
-
Termite digestomes as a potential source of symbiotic microbiota for lignocelluloses degradation: a review.Pak J Biol Sci. 2014 Aug;17(8):956-63. doi: 10.3923/pjbs.2014.956.963. Pak J Biol Sci. 2014. PMID: 26031014 Review.
Cited by
-
Purified fibers in chemically defined synthetic diets destabilize the gut microbiome of an omnivorous insect model.Front Microbiomes. 2024;3:1477521. doi: 10.3389/frmbi.2024.1477521. Epub 2024 Dec 11. Front Microbiomes. 2024. PMID: 40114931 Free PMC article.
-
Gut microbiota and quantitative traits divergence at different altitude of long-tailed dwarf hamsters, Cricetulus longicaudatus.Front Microbiol. 2025 Jan 24;15:1531629. doi: 10.3389/fmicb.2024.1531629. eCollection 2024. Front Microbiol. 2025. PMID: 39925881 Free PMC article.
-
Cultivating crayfish (Procambarus clarkii) significantly enhances the quantity and diversity of soil microorganisms: evidence from the comparison of rice-wheat and rice-crayfish rotation models.Front Microbiol. 2025 Feb 3;16:1528883. doi: 10.3389/fmicb.2025.1528883. eCollection 2025. Front Microbiol. 2025. PMID: 39963490 Free PMC article.
-
Metagenomics reveals fibre fermentation and AMR pathways in red grouse (Lagopus scotica) microbiota.BMC Microbiol. 2025 Aug 19;25(1):520. doi: 10.1186/s12866-025-04280-1. BMC Microbiol. 2025. PMID: 40830751 Free PMC article.
-
Characterization of two GH10 enzymes with ability to hydrolyze pretreated Sorghum bicolor bagasse.Appl Microbiol Biotechnol. 2025 Apr 28;109(1):104. doi: 10.1007/s00253-025-13484-4. Appl Microbiol Biotechnol. 2025. PMID: 40295346 Free PMC article.
References
-
- Ni J, Tokuda G. Lignocellulose-degrading enzymes from termites and their symbiotic microbiota. Biotechnol Adv. 2013;31:838–50. - PubMed
-
- Tokuda, G. Plant cell wall degradation in insects: recent progress on endogenous enzymes revealed by multi-omics technologies. In: Jurenka, R. editor. Advances in Insect Physiology. Cambridge: Academic Press; 2019. vol. 57. p. 97–136
-
- Brune A. Symbiotic digestion of lignocellulose in termite guts. Nat Rev Microbiol. 2014;12:168–80. - PubMed
-
- Brune A, Dietrich C. The gut microbiota of termites: digesting the diversity in the light of ecology and evolution. Annu Rev Microbiol. 2015;69:145–66. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
