

SARS-CoV-2-Specific T-Cell as a Potent Therapeutic Strategy against Immune Evasion of Emerging COVID-19 Variants
- PMID: 39408840
- PMCID: PMC11477143
- DOI: 10.3390/ijms251910512
SARS-CoV-2-Specific T-Cell as a Potent Therapeutic Strategy against Immune Evasion of Emerging COVID-19 Variants
Abstract
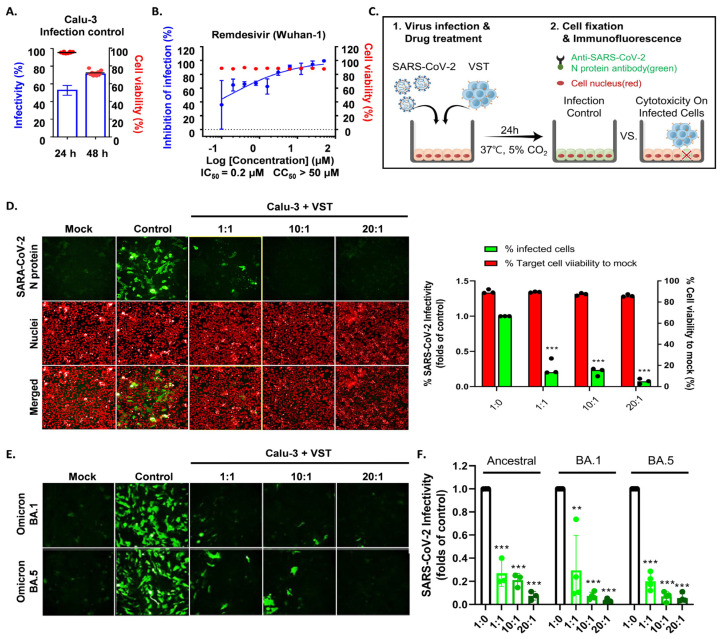
Despite advances in vaccination and therapies for coronavirus disease, challenges remain due to reduced antibody longevity and the emergence of virulent variants like Omicron (BA.1) and its subvariants (BA.1.1, BA.2, BA.3, and BA.5). This study explored the potential of adoptive immunotherapy and harnessing the protective abilities using virus-specific T cells (VSTs). Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) VSTs were generated by stimulating donor-derived peripheral blood mononuclear cells with spike, nucleocapsid, and membrane protein peptide mixtures. Phenotypic characterization, including T-cell receptor (TCR) vβ and pentamer analyses, was performed on the ex vivo-expanded cells. We infected human leukocyte antigen (HLA)-partially matched human Calu-3 cells with various authentic SARS-CoV-2 strains in a Biosafety Level 3 facility and co-cultured them with VSTs. VSTs exhibited a diverse TCR vβ repertoire, confirming their ability to target a broad range of SARS-CoV-2 antigens from both the ancestral and mutant strains, including Omicron BA.1 and BA.5. These ex vivo-expanded cells exhibited robust cytotoxicity and low alloreactivity against HLA-partially matched SARS-CoV-2-infected cells. Their cytotoxic effects were consistent across variants, targeting conserved spike and nucleocapsid epitopes. Our findings suggest that third-party partial HLA-matching VSTs could counter immune-escape mechanisms posed by emerging variants of concern.
Keywords: coronavirus disease; immunotherapy; severe acute respiratory syndrome coronavirus 2; viral immunity; virus-specific T cells.
Conflict of interest statement
Keon-Il Im, Nayoun Kim, Ui-Hyeon Oh, and Hye-Won Lee are employees and Seok-Goo Cho is the CEO of LucasBio, which develops products relevant to the subject matter discussed in this manuscript. The authors declare that this affiliation did not influence the design, execution, or interpretation of the research findings presented in this study. We have complied with the journal’s policy on competing interests, and all authors have disclosed any financial or personal relationships that could be perceived as potentially influencing the work.
Figures








References
-
- Rydyznski Moderbacher C., Ramirez S.I., Dan J.M., Grifoni A., Hastie K.M., Weiskopf D., Belanger S., Abbott R.K., Kim C., Choi J., et al. Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell. 2020;183:996–1012.e19. doi: 10.1016/j.cell.2020.09.038. - DOI - PMC - PubMed
-
- Lafon E., Diem G., Witting C., Zaderer V., Bellmann-Weiler R.M., Reindl M., Bauer A., Griesmacher A., Fux V., Hoermann G., et al. Potent SARS-CoV-2-specific T cell immunity and low anaphylatoxin levels correlate with mild disease progression in COVID-19 patients. Front. Immunol. 2021;12:684014. doi: 10.3389/fimmu.2021.684014. - DOI - PMC - PubMed
-
- Toniutto P., Falleti E., Cmet S., Cussigh A., Veneto L., Bitetto D., Fornasiere E., Fumolo E., Fabris C., Sartor A., et al. Past COVID-19 and immunosuppressive regimens affect the long-term response to anti-SARS-CoV-2 vaccination in liver transplant recipients. J. Hepatol. 2022;77:152–162. doi: 10.1016/j.jhep.2022.02.015. - DOI - PMC - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
- HI23C015100/Ministry of Health and Welfare, Republic of Korea
- NRF-2023M3A9G6057281 and NRF-2022M3A9J1081343/National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT)
- HI22C131400/a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
