

The eIF3a translational control axis in the Wnt/β-catenin signaling pathway and colon tumorigenesis
- PMID: 39413959
- PMCID: PMC11646415
- DOI: 10.1016/j.canlet.2024.217303
The eIF3a translational control axis in the Wnt/β-catenin signaling pathway and colon tumorigenesis
Abstract
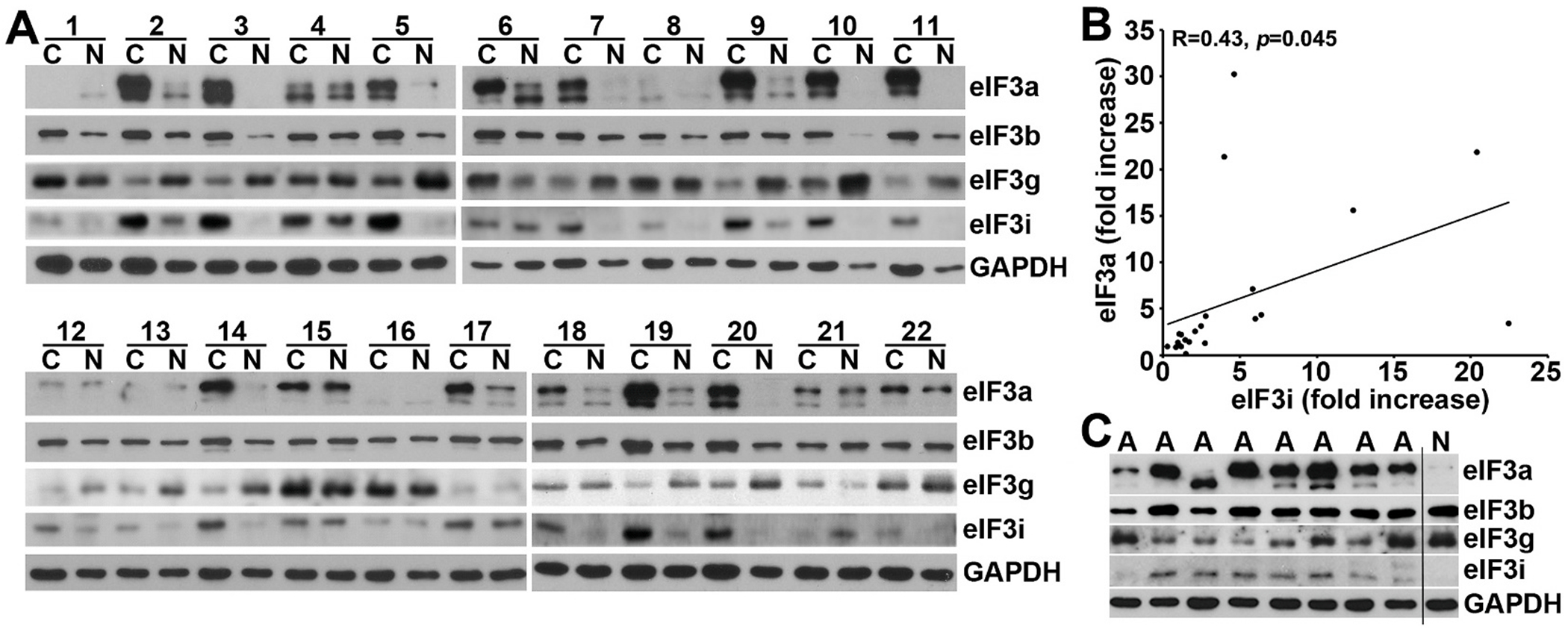
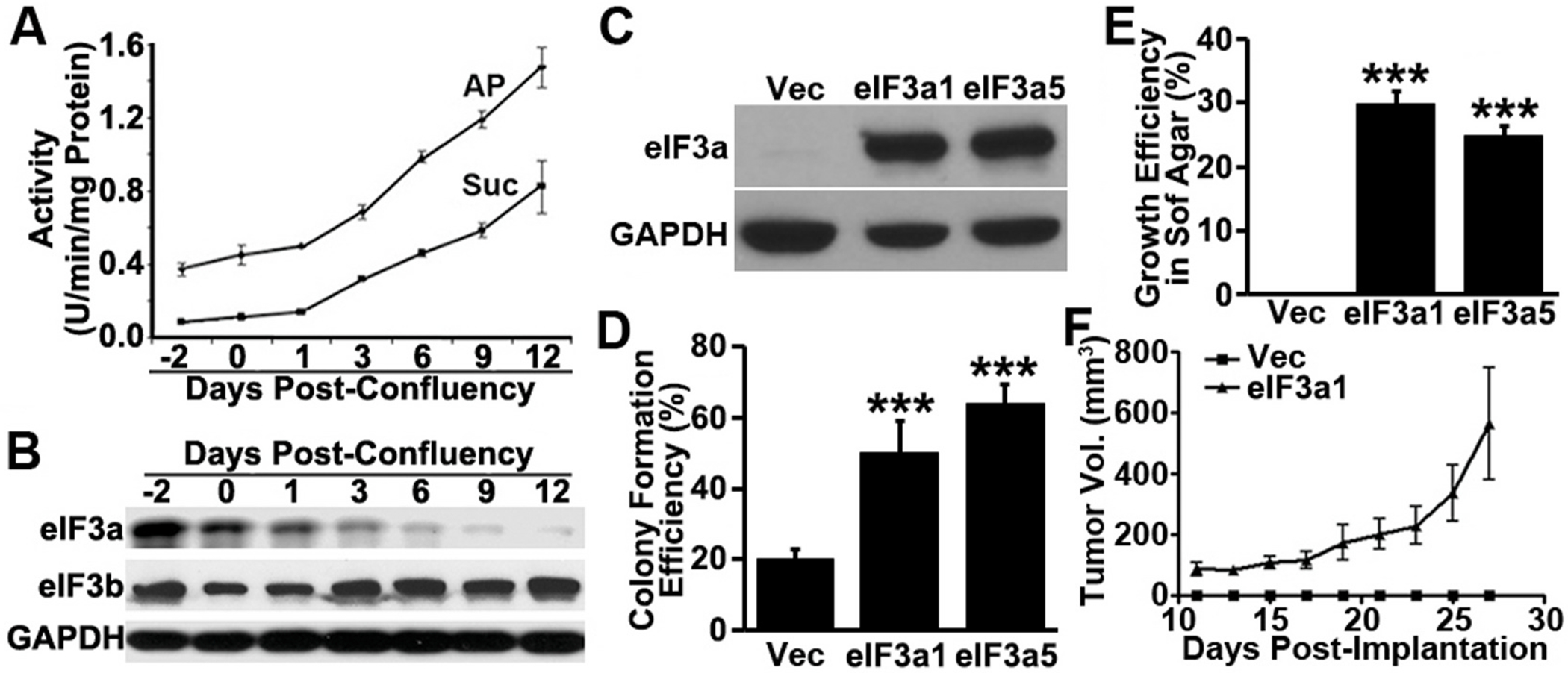
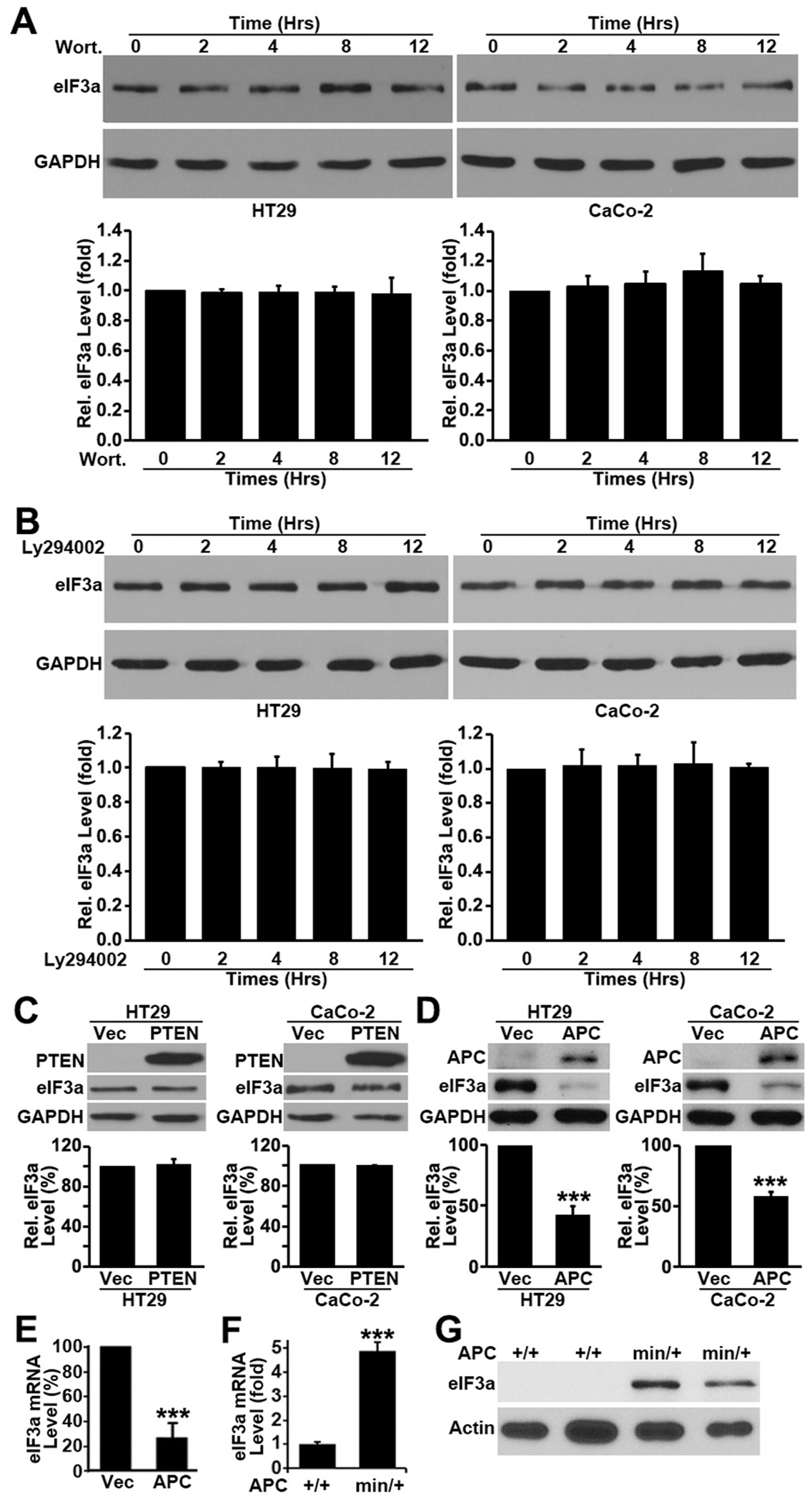
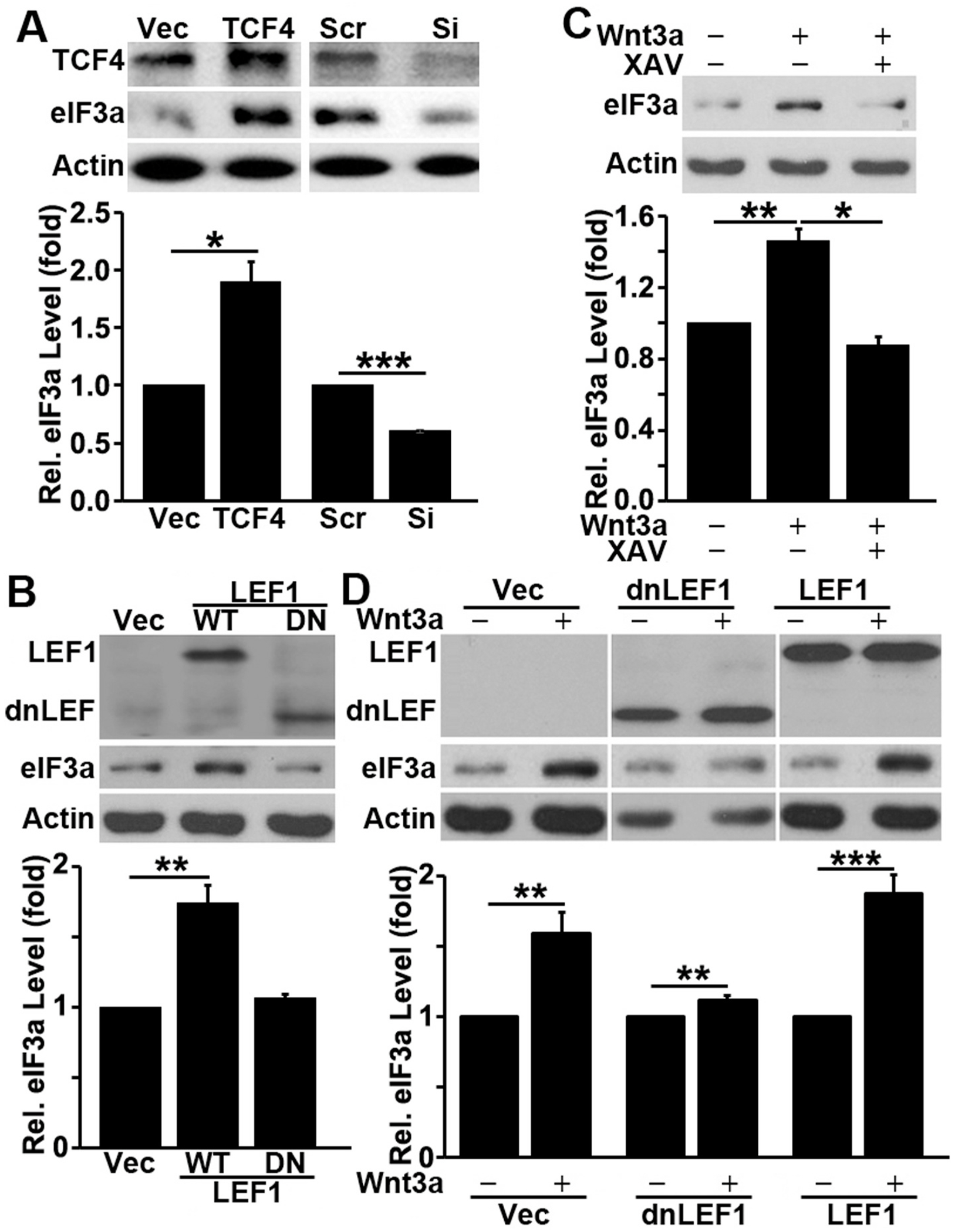
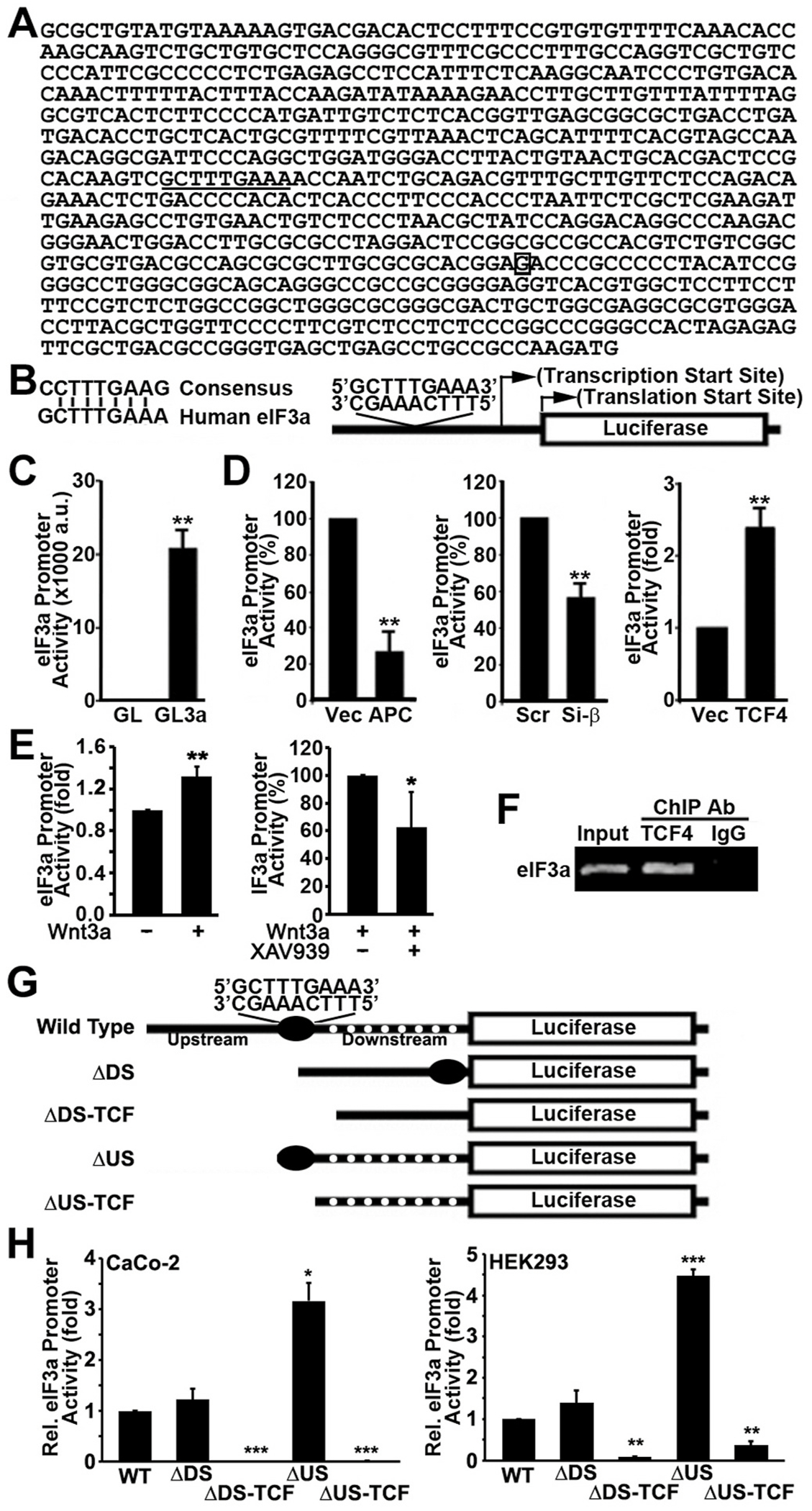
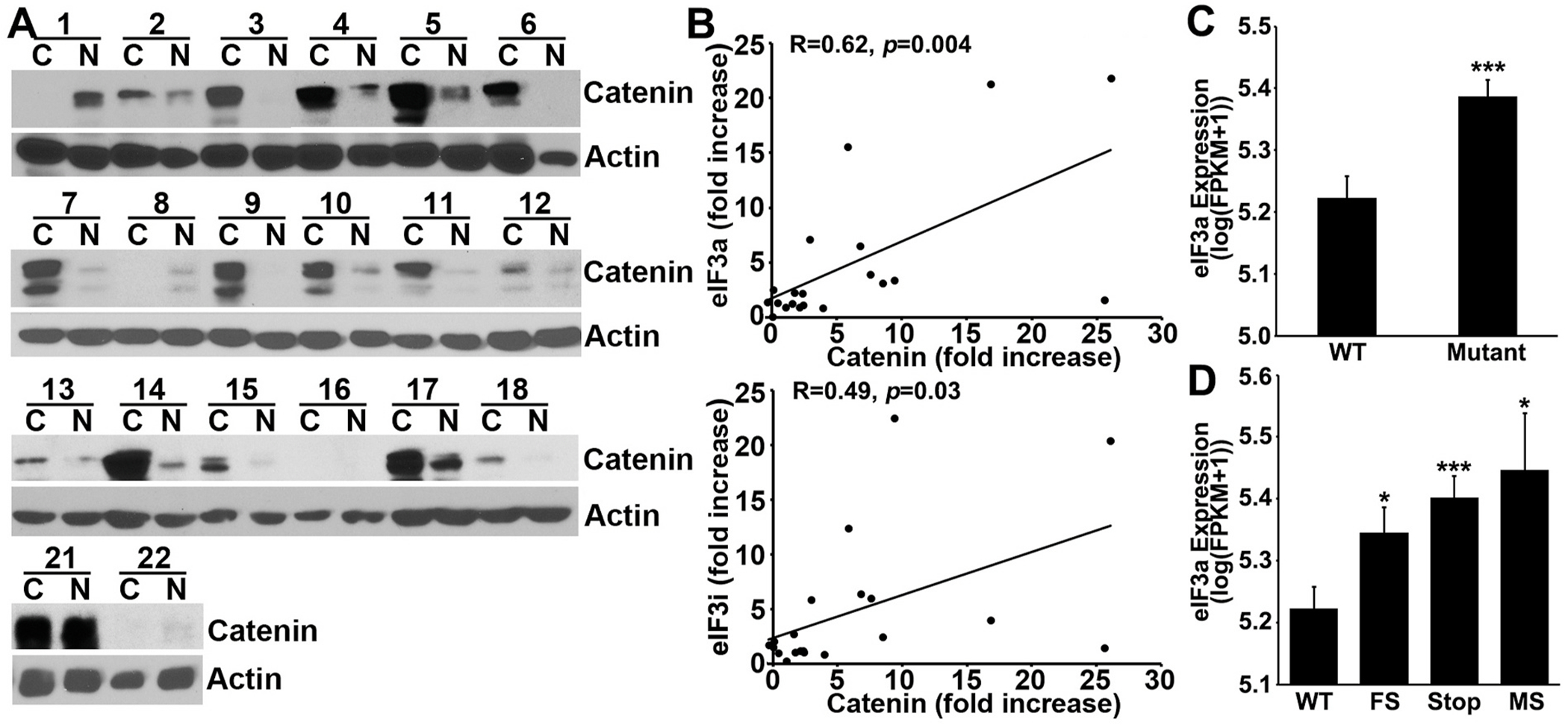
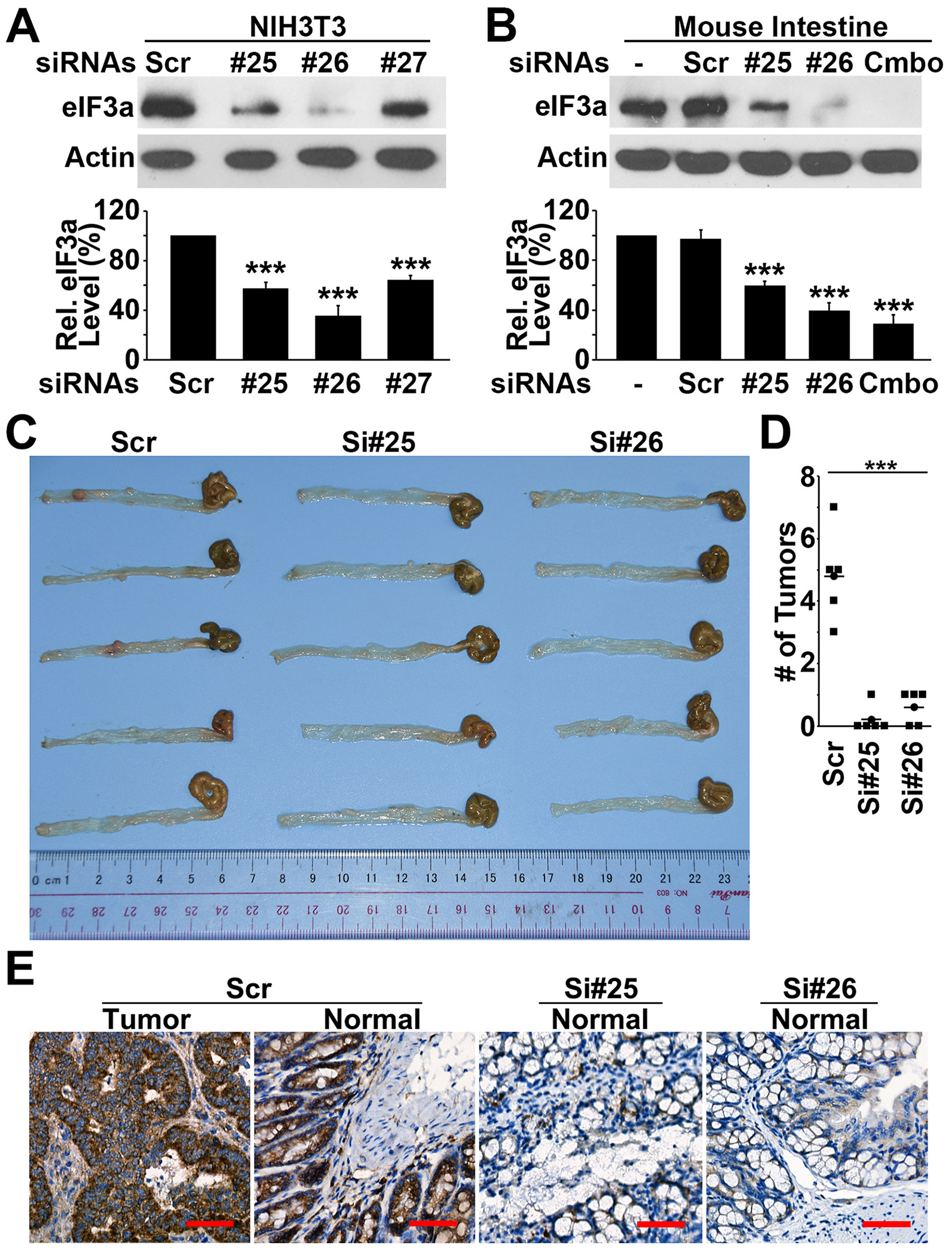
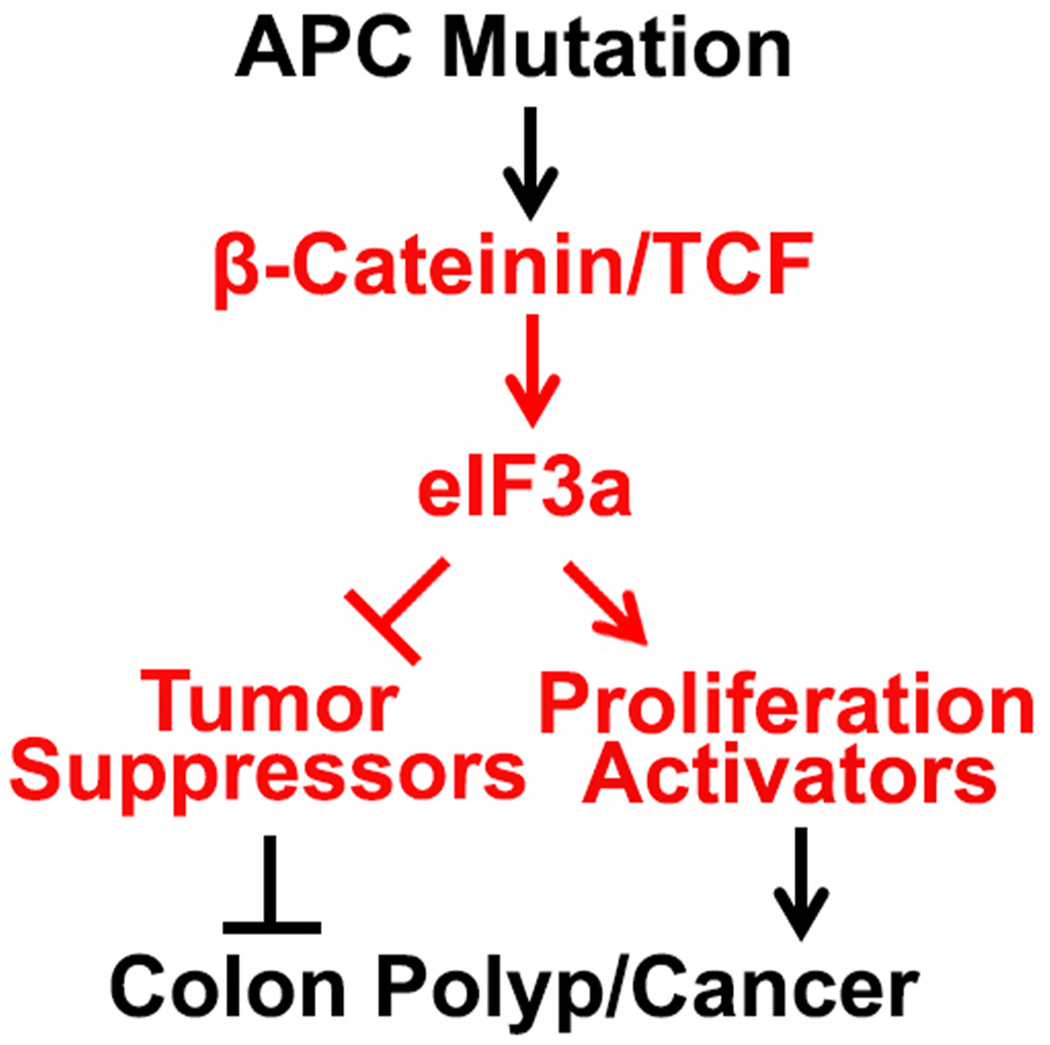
Translational initiation in protein synthesis is an important regulatory step in gene expression and its dysregulation may result in diseases such as cancer. Translational control by eIF4E/4E-BP has been well studied and contributes to mTOR signaling in various biological processes. Here, we report a novel translational control axis in the Wnt/β-catenin signaling pathway in colon tumorigenesis by eIF3a, a Yin-Yang factor in tumorigenesis and prognosis. We show that eIF3a expression is upregulated in human colon cancer tissues, pre-cancerous adenoma polyps, and associates with β-catenin level and APC mutation in human samples, and that eIF3a overexpression transforms intestinal epithelial cells. We also show that eIF3a expression is regulated by the Wnt/β-catenin signaling pathway with an active TCF/LEF binding site in its promoter and that eIF3a knockdown inhibits APC mutation-induced spontaneous colon tumorigenesis in APCmin/+ mice. Together, we conclude that eIF3a upregulation in colon cancer is due to APC mutation and it participates in colon tumorigenesis by adding a translational control axis in the Wnt/β-catenin signaling pathway and that it can serve as a potential target for colon cancer intervention.
Copyright © 2024 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Hershey JW, The role of eIF3 and its individual subunits in cancer, Biochim. Biophys. Acta 1849 (2015) 792–800. - PubMed
-
- Xu YC, Ruggero D, The role of translation control in tumorigenesis and its therapeutic implications, Annu Rev Cane Biol 4 (2020) 437–457.
-
- Dong Z, Zhang JT, Initiation factor eIF3 and regulation of mRNA translation, cell growth, and cancer, Crit. Rev. Oncol. Hematol 59 (2006) 169–180. - PubMed
-
- Ren D, Mo Y, Yang M, Wang D, Wang Y, Yan Q, Guo C, Xiong W, Wang F, Zeng Z, Emerging roles of tRNA in cancer, Cancer Lett. 563 (2023) 216170. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
