

Phosphorylation-driven epichaperome assembly is a regulator of cellular adaptability and proliferation
- PMID: 39414766
- PMCID: PMC11484706
- DOI: 10.1038/s41467-024-53178-5
Phosphorylation-driven epichaperome assembly is a regulator of cellular adaptability and proliferation
Abstract
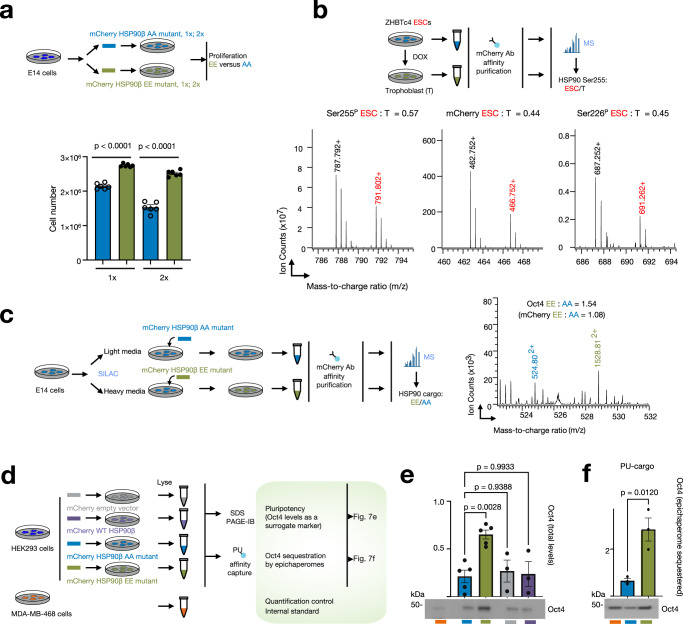
The intricate network of protein-chaperone interactions is crucial for maintaining cellular function. Recent discoveries have unveiled the existence of specialized chaperone assemblies, known as epichaperomes, which serve as scaffolding platforms that orchestrate the reconfiguration of protein-protein interaction networks, thereby enhancing cellular adaptability and proliferation. This study explores the structural and regulatory aspects of epichaperomes, with a particular focus on the role of post-translational modifications (PTMs) in their formation and function. A key finding is the identification of specific PTMs on HSP90, particularly at residues Ser226 and Ser255 within an intrinsically disordered region, as critical determinants of epichaperome assembly. Our data demonstrate that phosphorylation of these serine residues enhances HSP90's interactions with other chaperones and co-chaperones, creating a microenvironment conducive to epichaperome formation. Moreover, we establish a direct link between epichaperome function and cellular physiology, particularly in contexts where robust proliferation and adaptive behavior are essential, such as in cancer and pluripotent stem cell maintenance. These findings not only provide mechanistic insights but also hold promise for the development of novel therapeutic strategies targeting chaperone assemblies in diseases characterized by epichaperome dysregulation, thereby bridging the gap between fundamental research and precision medicine.
© 2024. The Author(s).
Conflict of interest statement
Memorial Sloan Kettering Cancer Center holds the intellectual rights to the epichaperome portfolio. G.Chiosis., A.R., and S.S. are inventors on the licensed intellectual property. All other authors declare no competing interests.
Figures









Update of
-
Phosphorylation-Driven Epichaperome Assembly: A Critical Regulator of Cellular Adaptability and Proliferation.Res Sq [Preprint]. 2024 Apr 3:rs.3.rs-4114038. doi: 10.21203/rs.3.rs-4114038/v1. Res Sq. 2024. Update in: Nat Commun. 2024 Oct 16;15(1):8912. doi: 10.1038/s41467-024-53178-5. PMID: 38645031 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
- R56 AG061869/AG/NIA NIH HHS/United States
- P20 GM113131/GM/NIGMS NIH HHS/United States
- R01 CA172546; P01 CA186866/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- R56 AG061869, R01 AG067598, R01 AG074004, R01 AG072599, R56 AG072599, RF1 AG071805/U.S. Department of Health & Human Services | NIH | National Institute on Aging (U.S. National Institute on Aging)
- R56 AG072599/AG/NIA NIH HHS/United States
- RF1 AG071805/AG/NIA NIH HHS/United States
- R01 AG074004/AG/NIA NIH HHS/United States
- R01 AG072599/AG/NIA NIH HHS/United States
- U01 AG032969/AG/NIA NIH HHS/United States
- R01 CA172546/CA/NCI NIH HHS/United States
- R01 AG085572/AG/NIA NIH HHS/United States
- R01 AG067598/AG/NIA NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- P01 CA186866/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
