

Zinc finger nuclease-mediated gene editing in hematopoietic stem cells results in reactivation of fetal hemoglobin in sickle cell disease
- PMID: 39414860
- PMCID: PMC11484757
- DOI: 10.1038/s41598-024-74716-7
Zinc finger nuclease-mediated gene editing in hematopoietic stem cells results in reactivation of fetal hemoglobin in sickle cell disease
Abstract
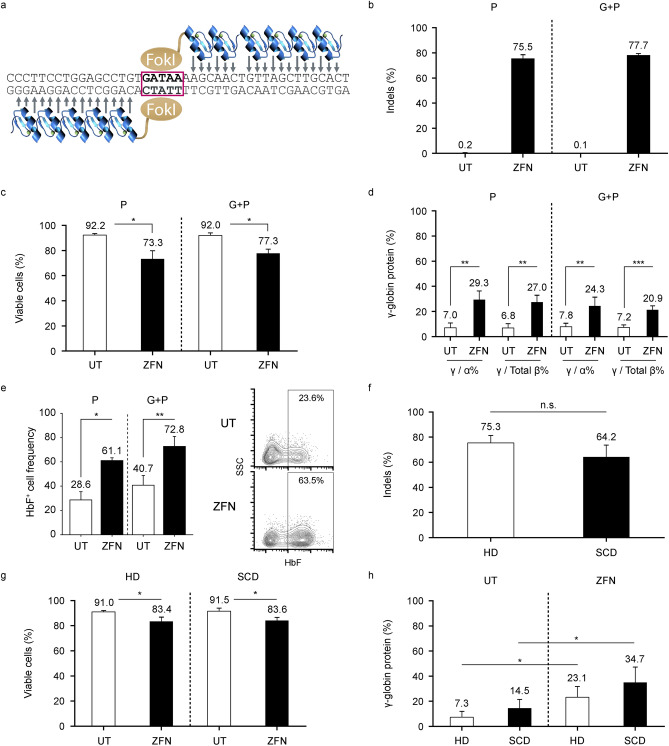
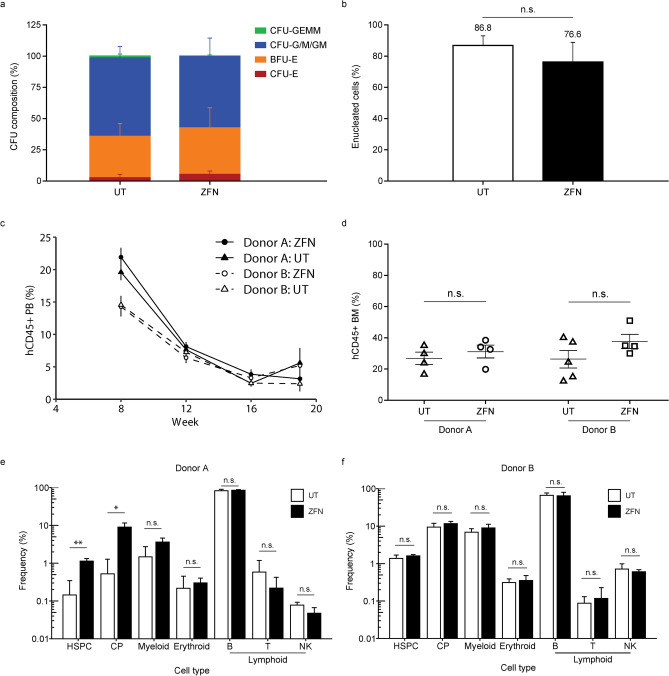
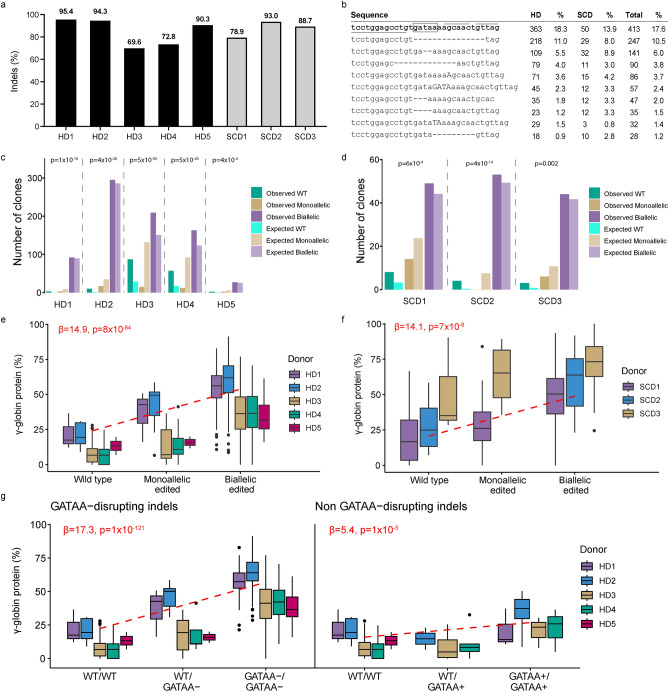
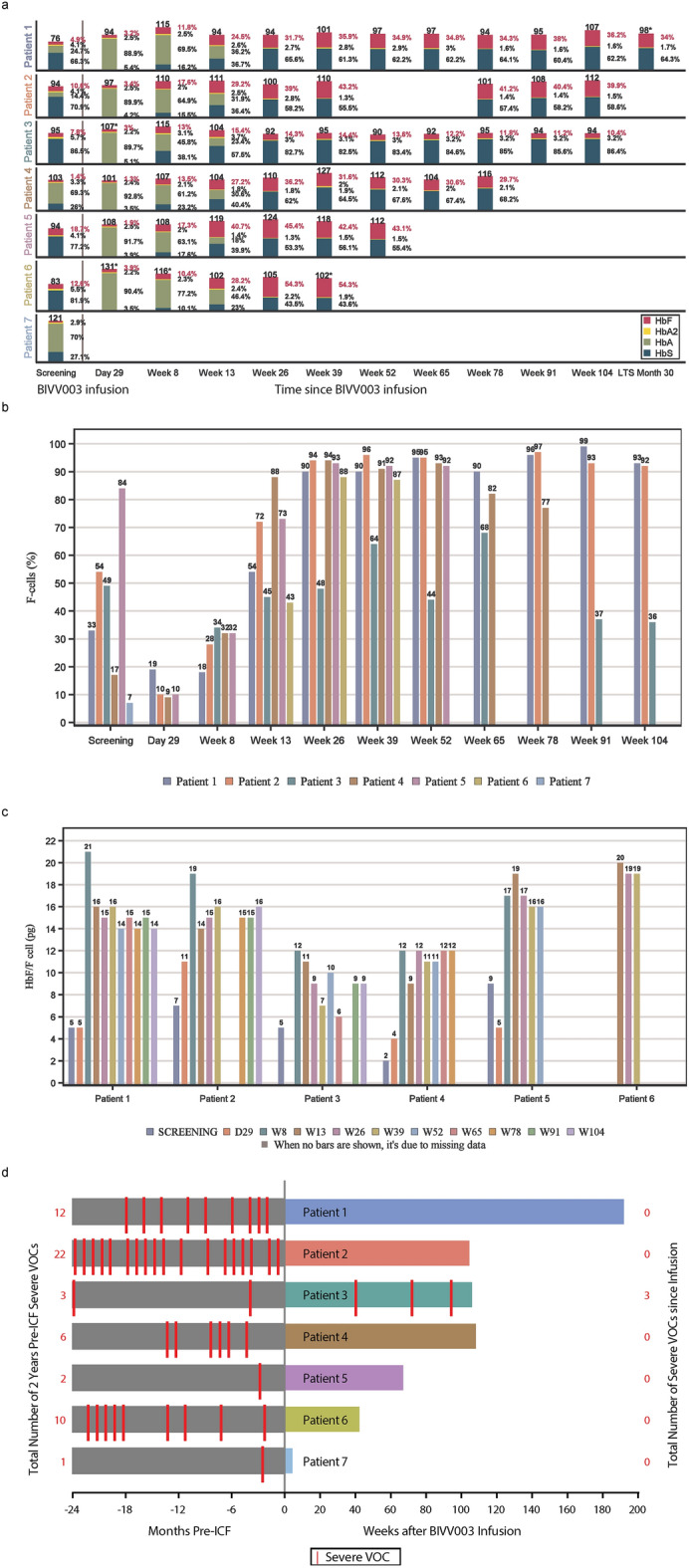
BIVV003 is a gene-edited autologous cell therapy in clinical development for the potential treatment of sickle cell disease (SCD). Hematopoietic stem cells (HSC) are genetically modified with mRNA encoding zinc finger nucleases (ZFN) that target and disrupt a specific regulatory GATAA motif in the BCL11A erythroid enhancer to reactivate fetal hemoglobin (HbF). We characterized ZFN-edited HSC from healthy donors and donors with SCD. Results of preclinical studies show that ZFN-mediated editing is highly efficient, with enriched biallelic editing and high frequency of on-target indels, producing HSC capable of long-term multilineage engraftment in vivo, and express HbF in erythroid progeny. Interim results from the Phase 1/2 PRECIZN-1 study demonstrated that BIVV003 was well-tolerated in seven participants with SCD, of whom five of the six with more than 3 months of follow-up displayed increased total hemoglobin and HbF, and no severe vaso-occlusive crises. Our data suggest BIVV003 represents a compelling and novel cell therapy for the potential treatment of SCD.
© 2024. The Author(s).
Conflict of interest statement
AH and SL are employees of Sanofi and may hold shares and/or stock options in the company. HL, KM, BV, DL, DR, PR, RP, TH, YL, GMR, IG and VH were employees of Sanofi at the time of the study. AR, MC, and BH are employees of Sangamo and may hold shares and/or stock options in the company. RB and BC were employees of Sangamo at the time of study. NU and JT have no conflict of interest. MA receives honorarium from Bristol Myers Squibb, hold stock from Cytodine Inc, and participates in speaker bureaus for Bristol Myers Squibb, Gilead, and Abbvie. MCW has served as consultant for Ensoma, Inc., Vertex pharmaceuticals, AllCells, Inc, and BioLabs, Inc.
Figures


References
-
- Hebbel, R. P., Osarogiagbon, R. & Kaul, D. The endothelial biology of sickle cell disease: inflammation and a chronic vasculopathy. Microcirculation11, 129–151 (2004). - PubMed
-
- Kato, G. J. et al. Sickle cell disease. Nat Rev Dis Primers4, 18010 (2018). - PubMed
-
- Lettre, G. & Bauer, D. E. Fetal haemoglobin in sickle-cell disease: from genetic epidemiology to new therapeutic strategies. Lancet387, 2554–2564 (2016). - PubMed
