

Structure of endothelin ETB receptor-Gi complex in a conformation stabilized by unique NPxxL motif
- PMID: 39414992
- PMCID: PMC11484851
- DOI: 10.1038/s42003-024-06905-z
Structure of endothelin ETB receptor-Gi complex in a conformation stabilized by unique NPxxL motif
Abstract
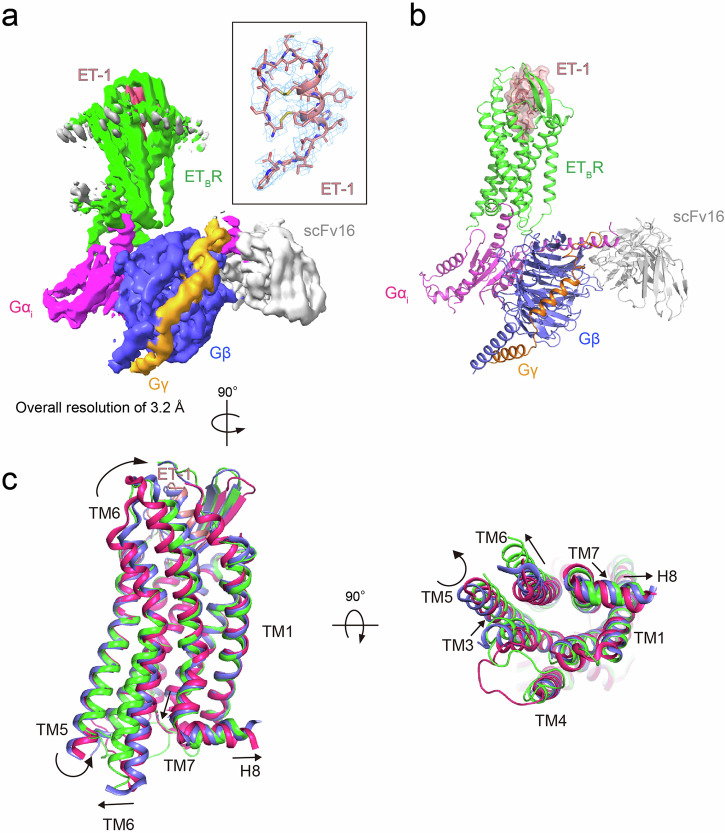
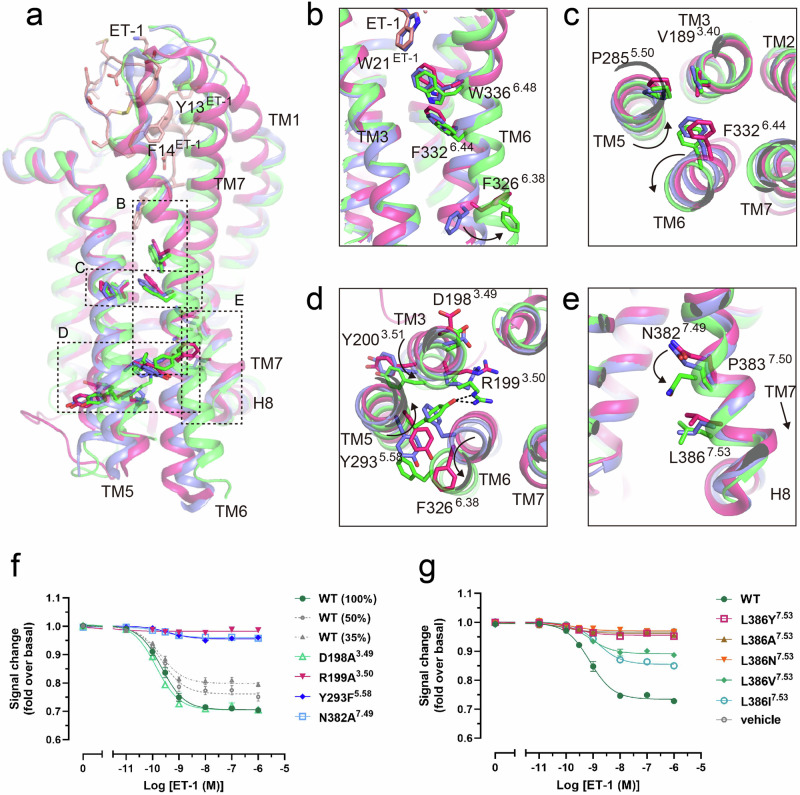
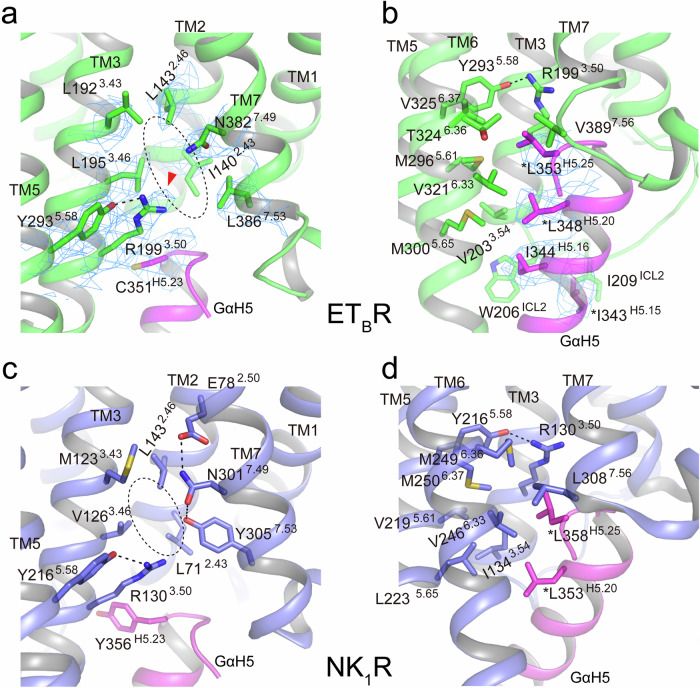
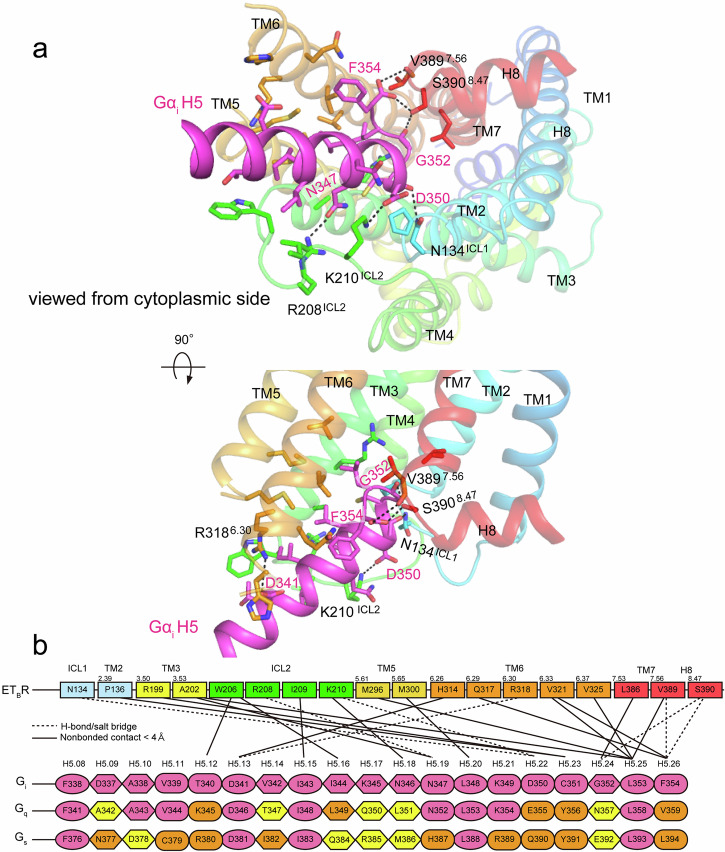
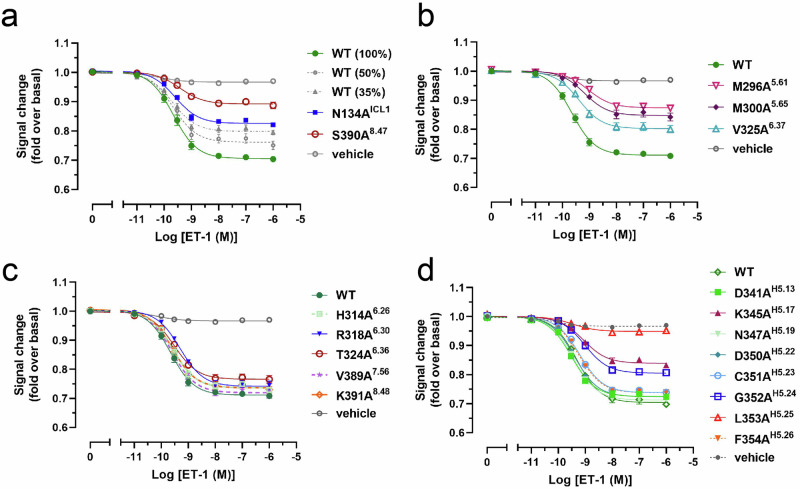
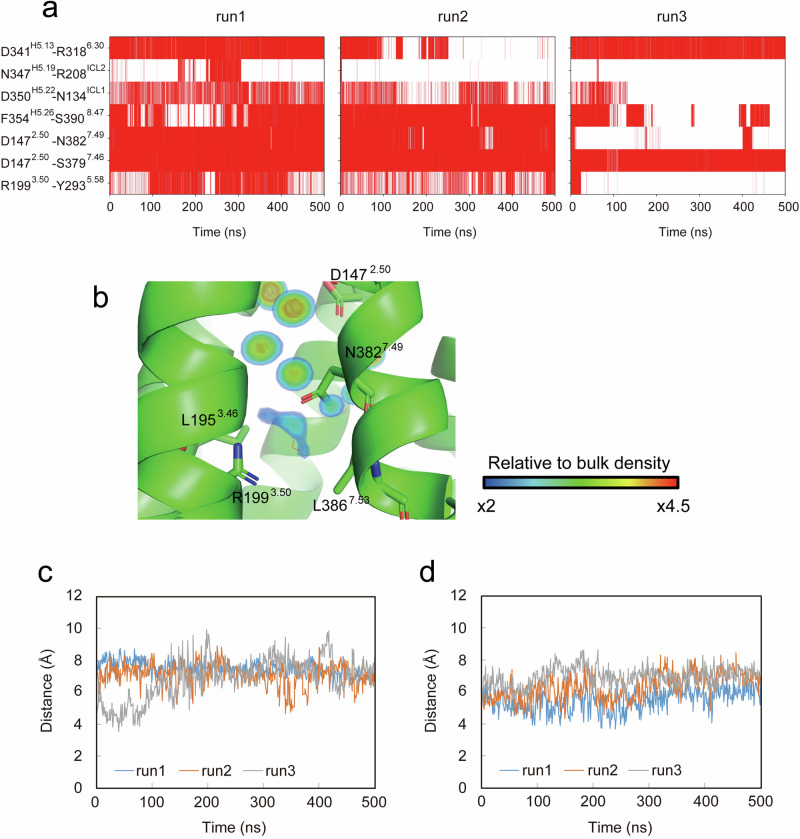
Endothelin type B receptor (ETBR) plays a crucial role in regulating blood pressure and humoral homeostasis, making it an important therapeutic target for related diseases. ETBR activation by the endogenous peptide hormones endothelin (ET)-1-3 stimulates several signaling pathways, including Gs, Gi/o, Gq/11, G12/13, and β-arrestin. Although the conserved NPxxY motif in transmembrane helix 7 (TM7) is important during GPCR activation, ETBR possesses the lesser known NPxxL motif. In this study, we present the cryo-EM structure of the ETBR-Gi complex, complemented by MD simulations and functional studies. These investigations reveal an unusual movement of TM7 to the intracellular side during ETBR activation and the essential roles of the diverse NPxxL motif in stabilizing the active conformation of ETBR and organizing the assembly of the binding pocket for the α5 helix of Gi protein. These findings enhance our understanding of the interactions between GPCRs and G proteins, thereby advancing the development of therapeutic strategies.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
