

This is a preprint.
Cardiolipin deficiency disrupts electron transport chain to drive steatohepatitis
- PMID: 39416056
- PMCID: PMC11482932
- DOI: 10.1101/2024.10.10.617517
Cardiolipin deficiency disrupts electron transport chain to drive steatohepatitis
Abstract
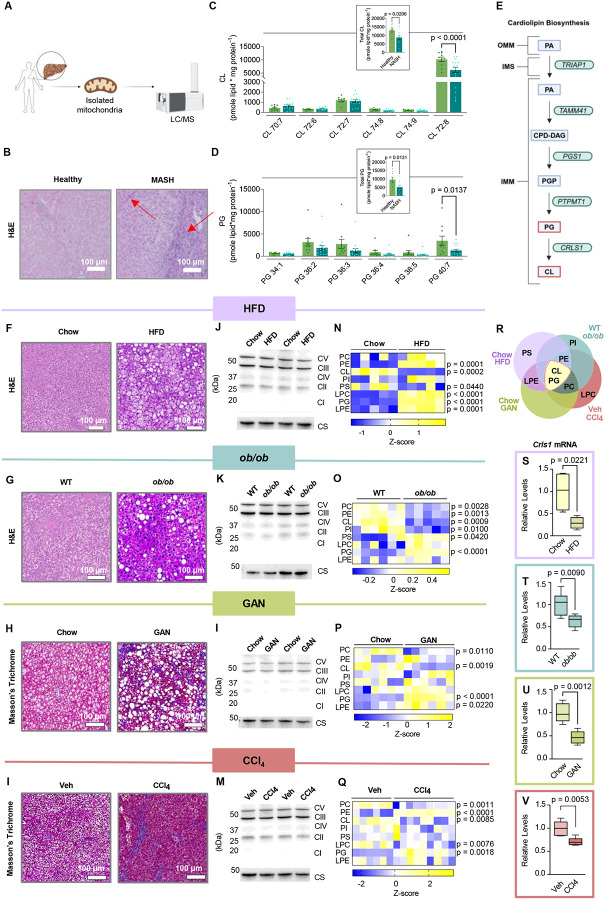
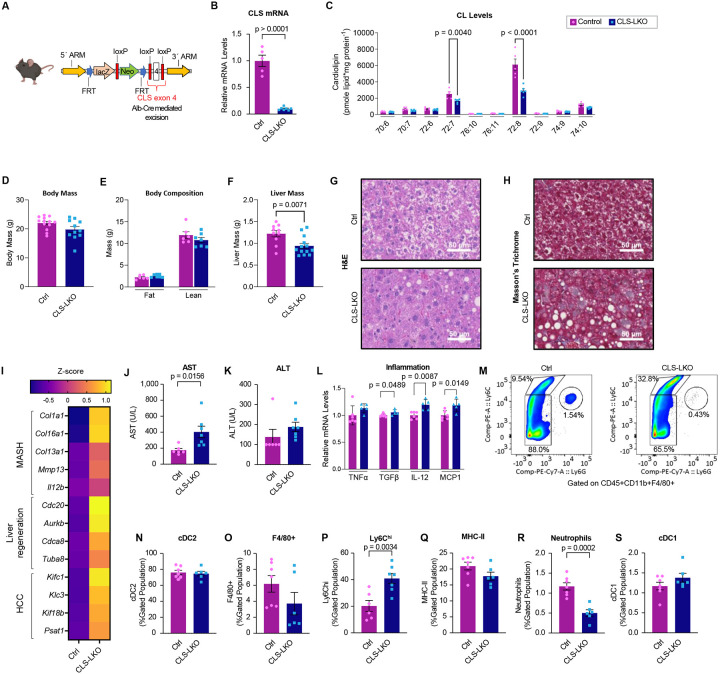
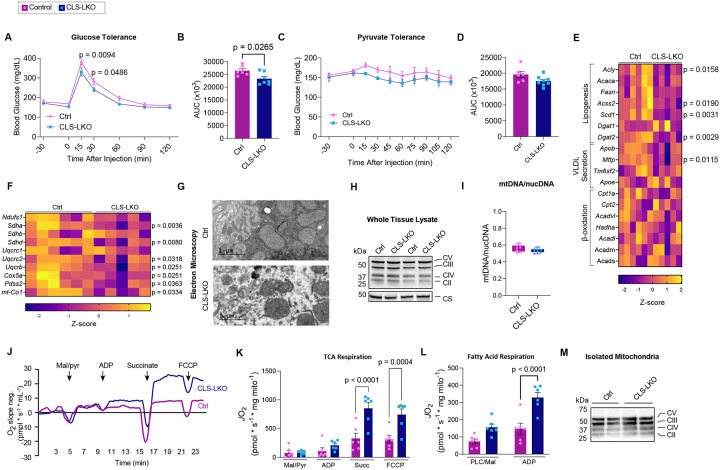
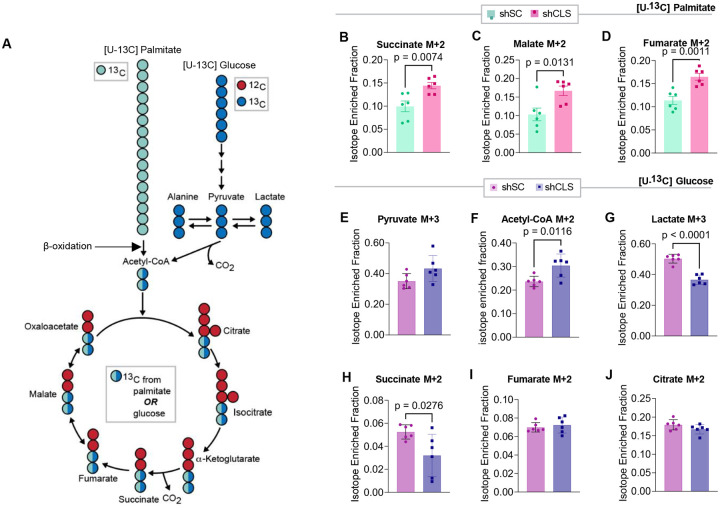
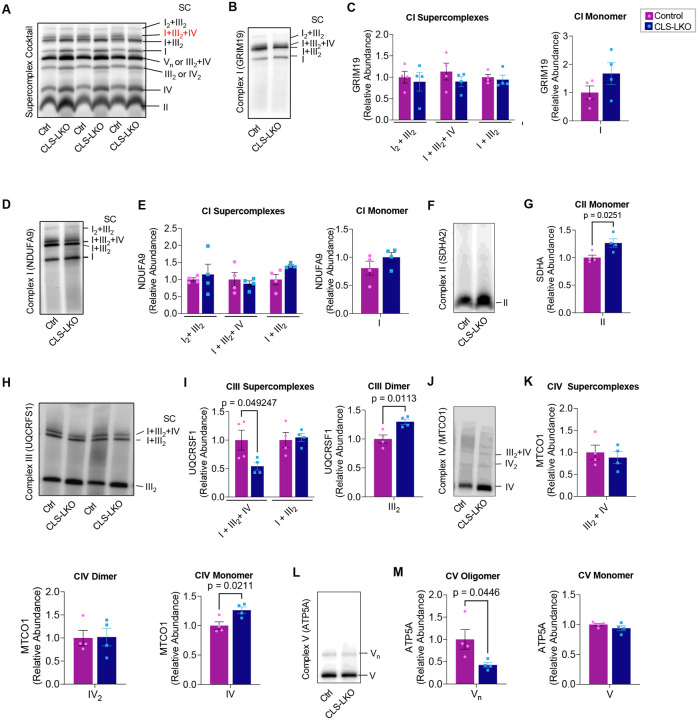
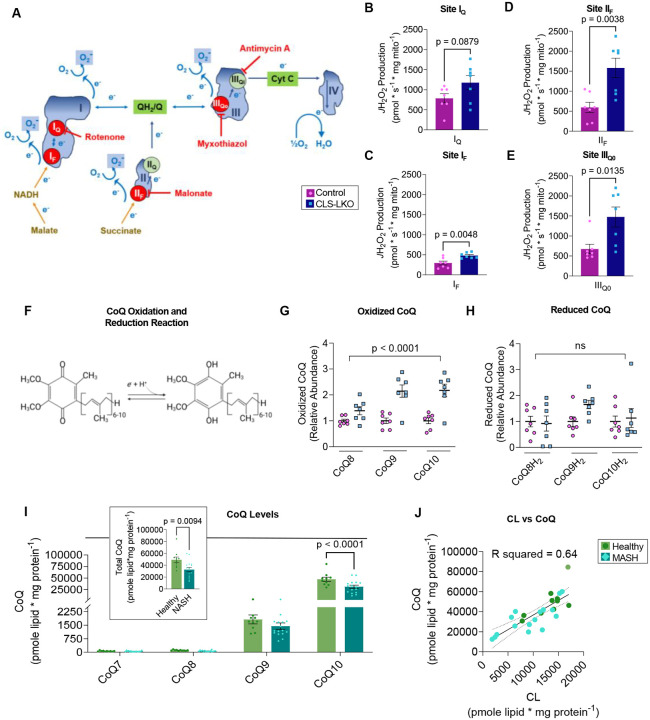
Metabolic dysfunction-associated steatotic liver disease (MASLD) is a progressive disorder marked by lipid accumulation, leading to metabolic dysfunction-associated steatohepatitis (MASH). A key feature of the transition to MASH involves oxidative stress resulting from defects in mitochondrial oxidative phosphorylation (OXPHOS). Here, we show that pathological alterations in the lipid composition of the inner mitochondrial membrane (IMM) directly instigate electron transfer inefficiency to promote oxidative stress. Specifically, mitochondrial cardiolipin (CL) was downregulated with MASLD/MASH in humans and in mice. Hepatocyte-specific CL synthase knockout (CLS-LKO) led to spontaneous and robust MASH with extensive steatotic and fibrotic phenotype. Loss of CL paradoxically increased mitochondrial respiratory capacity but also reduced the formation of I+III2+IV respiratory supercomplex, promoted electron leak primarily at sites IIIQO and IIF of the electron transport chain, and disrupted the propensity of coenzyme Q (CoQ) to become reduced. Thus, low mitochondrial CL disrupts electron transport chain to promote oxidative stress and contributes to pathogenesis of MASH.
Conflict of interest statement
Competing interests: the authors declare no competing interests.
Figures

References
-
- Younossi Z.M., et al. Global epidemiology of nonalcoholic fatty liver disease—Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 64, 73–84 (2015). - PubMed
-
- Miao L.T., Giovanni; Byrne, Christopher D; Cao, Ying-Ying; Zheng, Ming-Hua. Current status and future trends of the global burden of MASLD. Trends in Endocrinology & Metabolism 29, 1043–2760 (2024). - PubMed
-
- John B.S.S. Nonalcoholic Steatohepatitis (NASH). in StatPearls (StatPearls Publishing, Treasure Island, 2023). - PubMed
Publication types
Grants and funding
- S10 OD016232/OD/NIH HHS/United States
- F31 DK130555/DK/NIDDK NIH HHS/United States
- R01 DK128819/DK/NIDDK NIH HHS/United States
- UM1 TR004409/TR/NCATS NIH HHS/United States
- R01 GM144613/GM/NIGMS NIH HHS/United States
- R21 DK115991/DK/NIDDK NIH HHS/United States
- R01 DK127979/DK/NIDDK NIH HHS/United States
- R01 AG074535/AG/NIA NIH HHS/United States
- U01 CA272529/CA/NCI NIH HHS/United States
- R01 DK131609/DK/NIDDK NIH HHS/United States
- R35 GM151245/GM/NIGMS NIH HHS/United States
- R01 DK130296/DK/NIDDK NIH HHS/United States
- T32 DK091317/DK/NIDDK NIH HHS/United States
- S10 OD021505/OD/NIH HHS/United States
- R01 DK112826/DK/NIDDK NIH HHS/United States
- P30 CA042014/CA/NCI NIH HHS/United States
- R01 DK107397/DK/NIDDK NIH HHS/United States
- U54 DK110858/DK/NIDDK NIH HHS/United States
- F31 DK127603/DK/NIDDK NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- R01 HL170575/HL/NHLBI NIH HHS/United States
- R01 CA222570/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources