

Reverse hierarchical DED assembly in the cFLIP-procaspase-8 and cFLIP-procaspase-8-FADD complexes
- PMID: 39419969
- PMCID: PMC11487272
- DOI: 10.1038/s41467-024-53306-1
Reverse hierarchical DED assembly in the cFLIP-procaspase-8 and cFLIP-procaspase-8-FADD complexes
Erratum in
-
Publisher Correction: Reverse hierarchical DED assembly in the cFLIP-procaspase-8 and cFLIP-procaspase-8-FADD complexes.Nat Commun. 2024 Nov 6;15(1):9584. doi: 10.1038/s41467-024-54028-0. Nat Commun. 2024. PMID: 39505876 Free PMC article. No abstract available.
Abstract
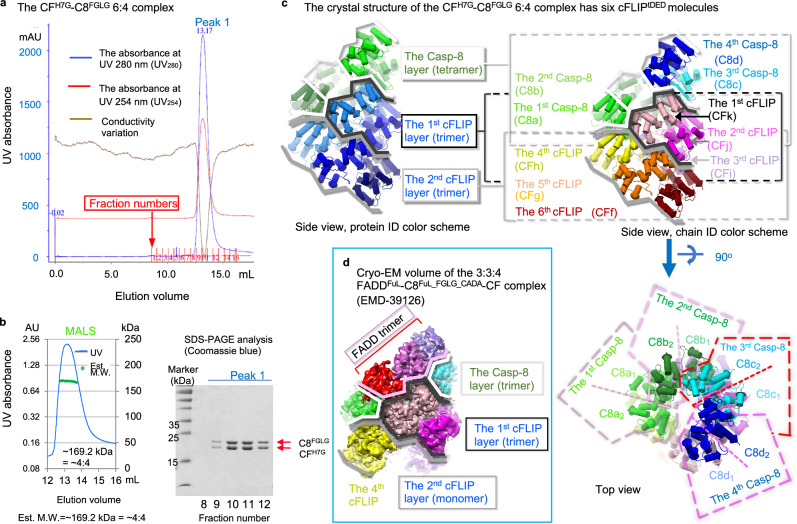
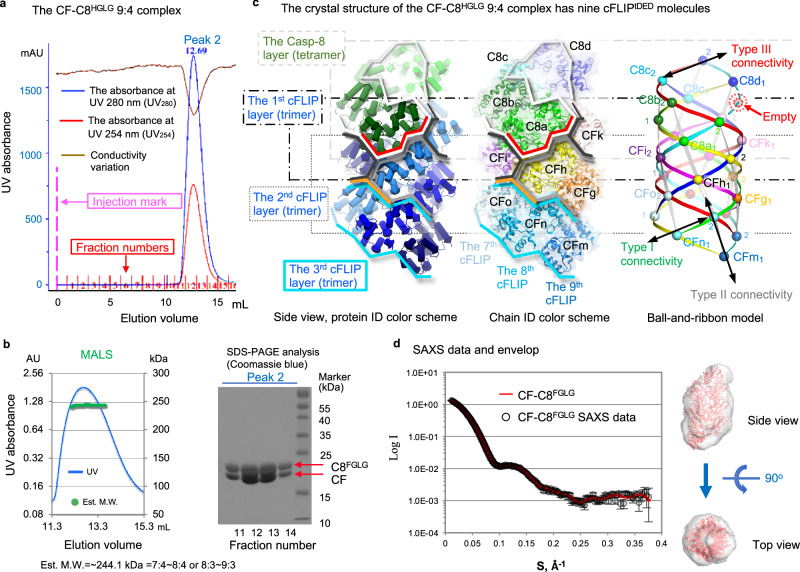
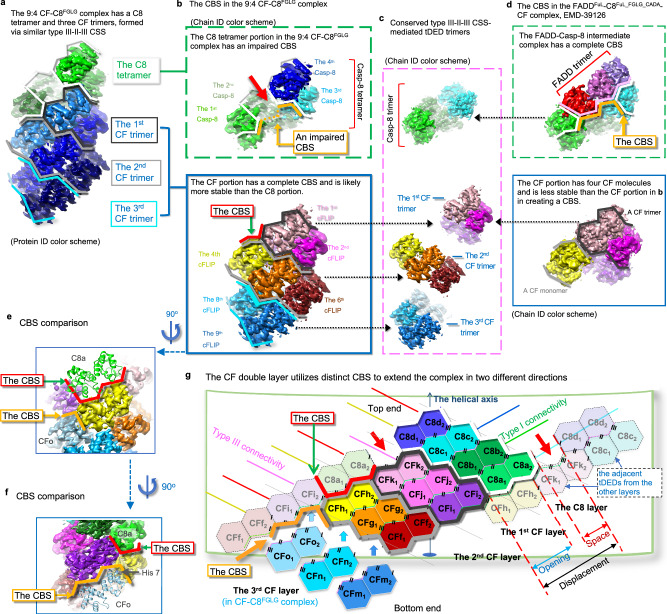
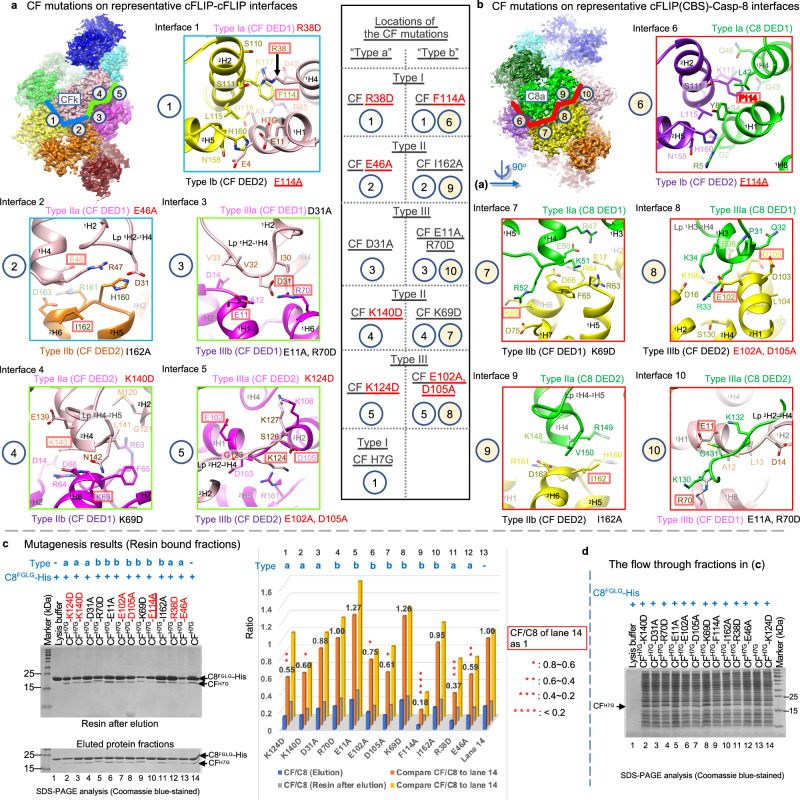
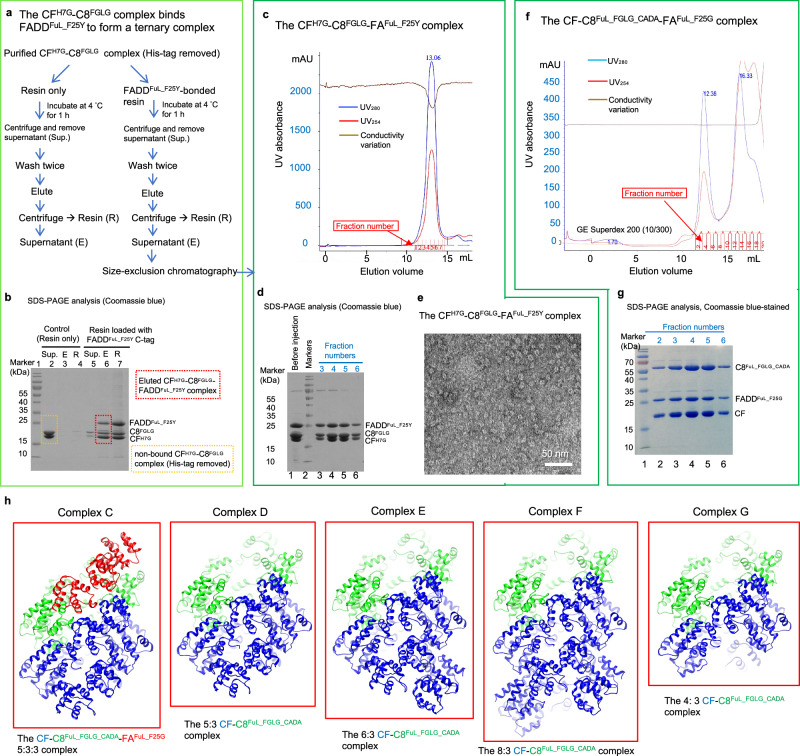
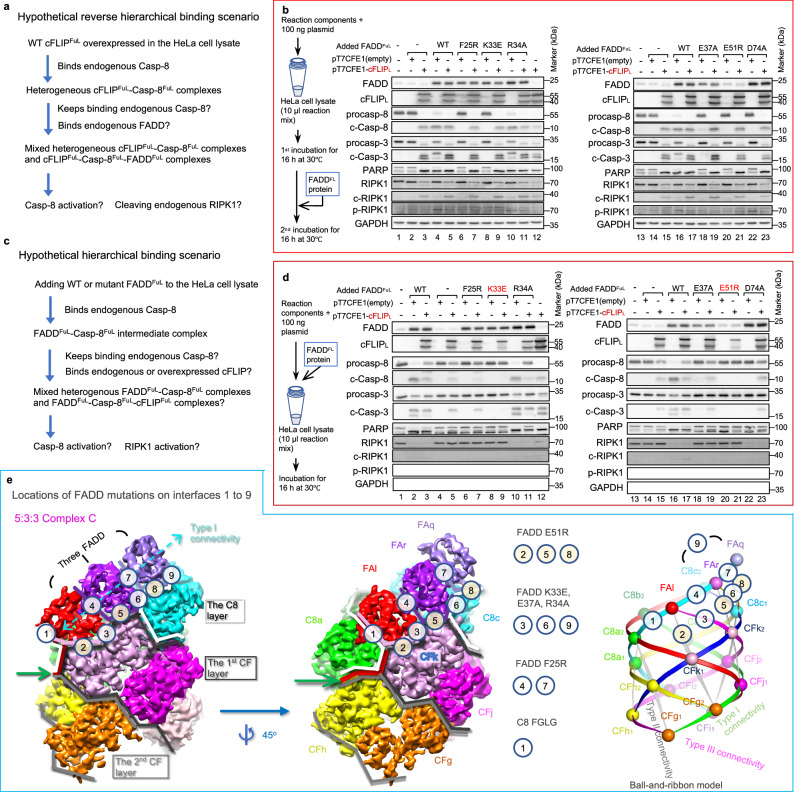
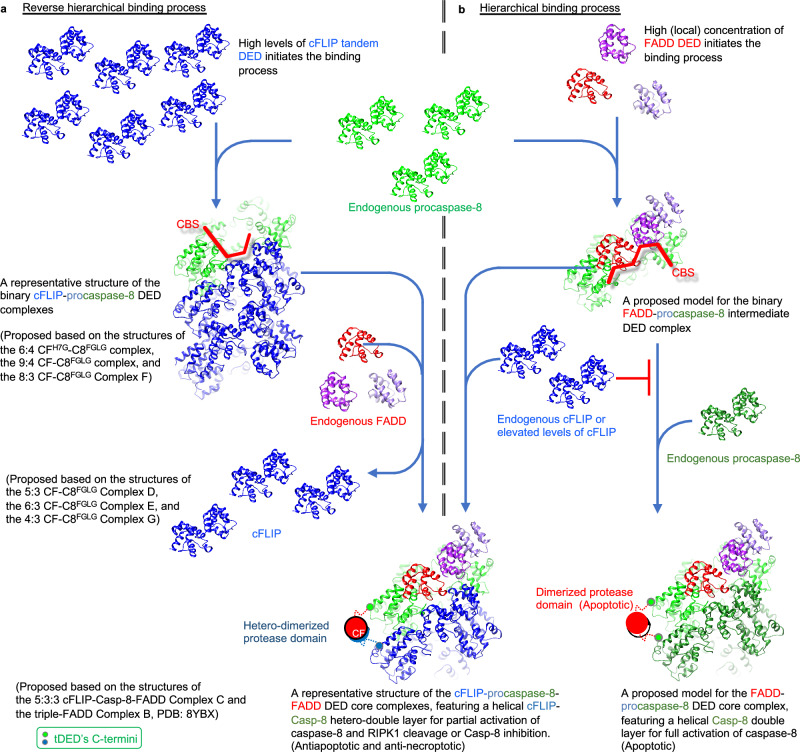
cFLIP, a master anti-apoptotic regulator, targets the FADD-induced DED complexes of procaspase-8 in death receptor and ripoptosome signaling pathways. Several tumor cells maintain relatively high levels of cFLIP in achieving their immortality. However, understanding the three-dimensional regulatory mechanism initiated or mediated by elevated levels of cFLIP has been limited by the absence of the atomic coordinates for cFLIP-induced DED complexes. Here we report the crystal plus cryo-EM structures to uncover an unconventional mechanism where cFLIP and procaspase-8 autonomously form a binary tandem DED complex, independent of FADD. This complex gains the ability to recruit FADD, thereby allosterically modulating cFLIP assembly and partially activating caspase-8 for RIPK1 cleavage. Our structure-guided mutagenesis experiments provide critical insights into these regulatory mechanisms, elucidating the resistance to apoptosis and necroptosis in achieving immortality. Finally, this research offers a unified model for the intricate bidirectional hierarchy-based processes using multiprotein helical assembly to govern cell fate decisions.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
- AS-TP-107-L16/Academia Sinica
- AS-102-TP-B14-2/Academia Sinica
- AS-TP-107-L16-1/Academia Sinica
- AS-102-TP-B14/Academia Sinica
- AS-TP-107-L16-2/Academia Sinica
- AS-102-TP-B14-1/Academia Sinica
- AS-TP-107-L16-3/Academia Sinica
- 110-2311-B-001-015-/Ministry of Science and Technology, Taiwan (Ministry of Science and Technology of Taiwan)
- 109-2311-B-001-016-/Ministry of Science and Technology, Taiwan (Ministry of Science and Technology of Taiwan)
- 108-2311-B-001-018-/Ministry of Science and Technology, Taiwan (Ministry of Science and Technology of Taiwan)
- 107-2320-B-001-018-/Ministry of Science and Technology, Taiwan (Ministry of Science and Technology of Taiwan)
- 107-2320-B-006-062-MY3/Ministry of Science and Technology, Taiwan (Ministry of Science and Technology of Taiwan)
- 111-2311-B-006-005-MY3/Ministry of Science and Technology, Taiwan (Ministry of Science and Technology of Taiwan)
- 111-2311-B-006-005-MY3/Ministry of Science and Technology, Taiwan (Ministry of Science and Technology of Taiwan)
LinkOut - more resources
Full Text Sources
Miscellaneous
