

G6PC3 is involved in spermatogenesis by maintaining meiotic sex chromosome inactivation
- PMID: 39420835
- PMCID: PMC11868919
- DOI: 10.3724/abbs.2024172
G6PC3 is involved in spermatogenesis by maintaining meiotic sex chromosome inactivation
Abstract
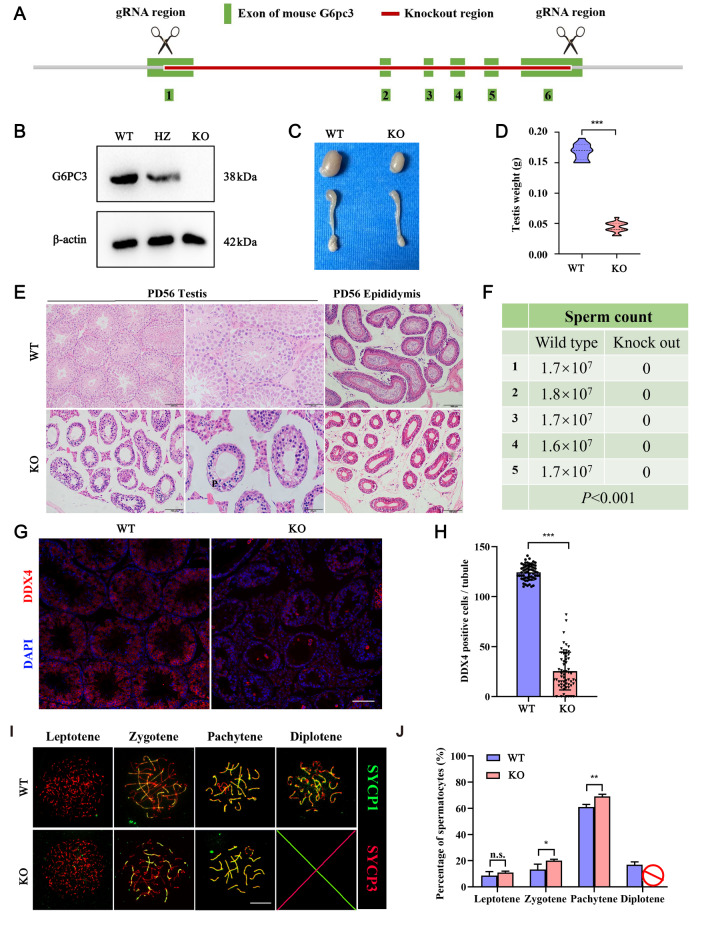
Meiosis, a process unique to germ cells, involves formation and repair of double-stranded nicks in DNA, pairing and segregation of homologous chromosomes, which ultimately achieves recombination of homologous chromosomes. Genetic abnormalities resulted from defects in meiosis are leading causes of infertility in humans. Meiotic sex chromosome inactivation (MSCI) plays a crucial role in the development of male germ cells in mammals, yet its underlying mechanisms remain poorly understood. In this study, we illustrate the predominant presence of a protein known as glucose 6 phosphatase catalyzed 3 (G6PC3) in pachytene spermatocytes, with a high concentration in the sex body (XY body), suggesting its significant involvement in male germ cell development. By employing CRISPR-Cas9 technology, we generate mice deficient in the G6pc3 gene, resulting in complete meiotic arrest at the pachytene stage in spermatocytes and are completely sterile. Additionally, we observe abnormal XY body formation and impaired MSCI in G6pc3-knockout spermatocytes. These findings underscore G6pc3 as a new essential regulator that is essential for meiotic progression. G6PC3 is involved in spermatocyte during male spermatogenesis development by the maintenance of meiosis chromosome silencing.
Keywords: glucose 6 phosphatase catalytic 3 (G6PC3); meiotic sex chromosome inactivation; pachytene arrest; sex chromosomes; spermatogenesis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Farini D, De Felici M. The beginning of meiosis in mammalian female germ cells: a never-ending story of intrinsic and extrinsic factors. Int J Mol Sci. . 2022;23:12571. doi: 10.3390/ijms232012571. - DOI - PMC - PubMed
-
- MacKenzie A, Vicory V, Lacefield S, Lichten M. Meiotic cells escape prolonged spindle checkpoint activity through kinetochore silencing and slippage. PLoS Genet. . 2023;19:e1010707. doi: 10.1371/journal.pgen.1010707. - DOI - PMC - PubMed
-
- Alavattam KG, Maezawa S, Andreassen PR, Namekawa SH. Meiotic sex chromosome inactivation and the XY body: a phase separation hypothesis. Cell Mol Life Sci. . 2021;79:18. doi: 10.1007/s00018-021-04075-3. - DOI - PMC - PubMed
-
- Abe H, Yeh YH, Munakata Y, Ishiguro KI, Andreassen PR, Namekawa SH. Active DNA damage response signaling initiates and maintains meiotic sex chromosome inactivation. Nat Commun. . 2022;13:7212. doi: 10.1038/s41467-022-34295-5. - DOI - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
