

A broadly reactive ultralong bovine antibody that can determine the integrity of foot-and-mouth disease virus capsids
- PMID: 39422666
- PMCID: PMC11488517
- DOI: 10.1099/jgv.0.002032
A broadly reactive ultralong bovine antibody that can determine the integrity of foot-and-mouth disease virus capsids
Abstract
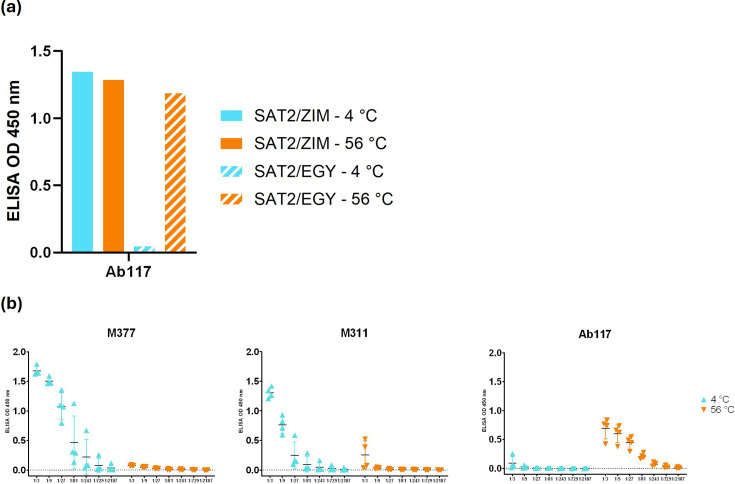
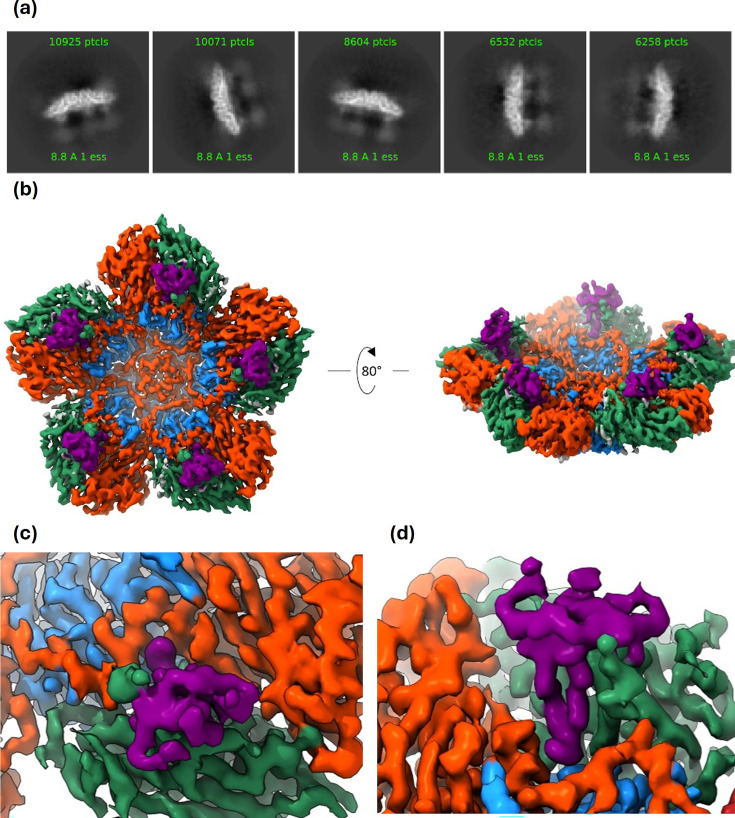
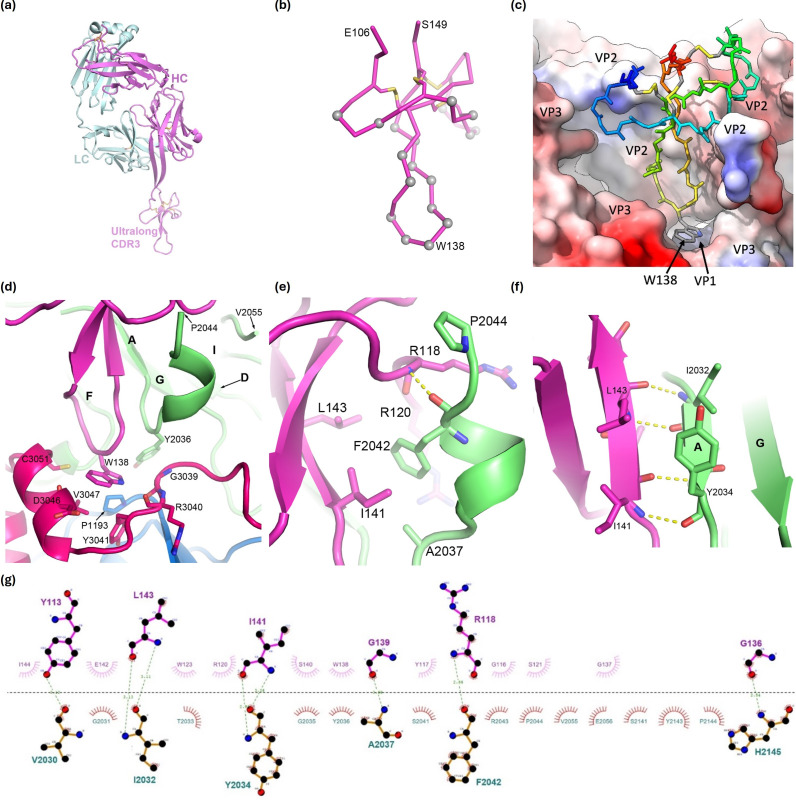
Foot-and-mouth disease vaccination using inactivated virus is suboptimal, as the icosahedral viral capsids often disassemble into antigenically distinct pentameric units during long-term storage, or exposure to elevated temperature or lowered pH, and thus raise a response that is no longer protective. Furthermore, as foot-and-mouth disease virus (FMDV)'s seven serotypes are antigenically diverse, cross-protection from a single serotype vaccine is limited, and most existing mouse and bovine antibodies and camelid single-domain heavy chain-only antibodies are serotype-specific. For quality control purposes, there is a real need for pan-serotype antibodies that clearly distinguish between pentamer (12S) and protective intact FMDV capsid. To date, few cross-serotype bovine-derived antibodies have been reported in the literature. We identify a bovine antibody with an ultralong CDR-H3, Ab117, whose structural analysis reveals that it binds to a deep, hydrophobic pocket on the interior surface of the capsid via the CDR-H3. Main-chain and hydrophobic interactions provide broad serotype specificity. ELISA analysis confirms that Ab117 is a novel pan-serotype and conformational epitope-specific 12S reagent, suitable for assessing capsid integrity.
Keywords: FMDV; pan-specific; single particle analysis; ultralong CDR antibody; vaccine quality assurance.
Conflict of interest statement
Some of the work presented in this study was presented within John D. Clarke’s doctoral thesis. The authors declare that no other conflicts of interest, direct or contrived, were identified in relation to this work. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
References
-
- The National Audit Office The 2001 Outbreak of Foot and Mouth Disease. Report. London: The National Audit Office, The Comptroller and Auditor General. 2002 102916454.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources