

Resource sharing of an infant gut microbiota synthetic community in combinations of human milk oligosaccharides
- PMID: 39423288
- PMCID: PMC11542058
- DOI: 10.1093/ismejo/wrae209
Resource sharing of an infant gut microbiota synthetic community in combinations of human milk oligosaccharides
Abstract
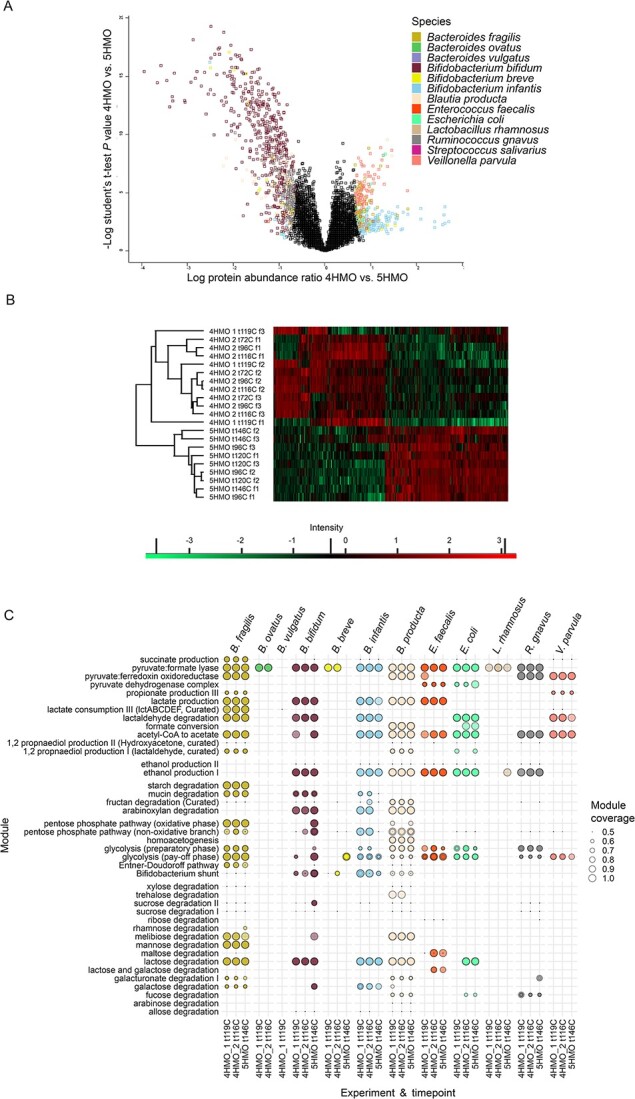
Quickly after birth, the gut microbiota is shaped via species acquisition and resource pressure. Breastmilk, and more specifically, human milk oligosaccharides are a determining factor in the formation of microbial communities and the interactions between bacteria. Prominent human milk oligosaccharide degraders have been rigorously characterized, but it is not known how the gut microbiota is shaped as a complex community. Here, we designed BIG-Syc, a synthetic community of 13 strains from the gut of vaginally born, breastfed infants. BIG-Syc replicated key compositional, metabolic, and proteomic characteristics of the gut microbiota of infants. Upon fermentation of a four and five human milk oligosaccharide mix, BIG-Syc demonstrated different compositional and proteomic profiles, with Bifidobacterium infantis and Bifidobacterium bifidum suppressing one another. The mix of five human milk oligosaccharides resulted in a more diverse composition with dominance of B. bifidum, whereas that with four human milk oligosaccharides supported the dominance of B. infantis, in four of six replicates. Reintroduction of bifidobacteria to BIG-Syc led to their engraftment and establishment of their niche. Based on proteomics and genome-scale metabolic models, we reconstructed the carbon source utilization and metabolite and gas production per strain. BIG-Syc demonstrated teamwork as cross-feeders utilized simpler carbohydrates, organic acids, and gases released from human milk oligosaccharide degraders. Collectively, our results showed that human milk oligosaccharides prompt resource-sharing for their complete degradation while leading to a different compositional and functional profile in the community. At the same time, BIG-Syc proved to be an accurate model for the representation of intra-microbe interactions.
Keywords: bifidobacteria; community dynamics; genome-scale metabolic modelling; human milk oligosaccharides; infant gut microbiota; synthetic community.
© The Author(s) 2024. Published by Oxford University Press on behalf of the International Society for Microbial Ecology.
Conflict of interest statement
JK, BB, and MM are employees of Danone Nutricia Research.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
-
- Urashima T, Hirabayashi J, Sato Set al. . Human milk oligosaccharides as essential tools for basic and application studies on galectins. Trends Glycosci Gly 2017;30:SE51–65. 10.4052/tigg.1734.1SE - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
