

Sirtuin 1 regulates the phenotype and functions of dendritic cells through Ido1 pathway in obesity
- PMID: 39424786
- PMCID: PMC11489582
- DOI: 10.1038/s41419-024-07125-3
Sirtuin 1 regulates the phenotype and functions of dendritic cells through Ido1 pathway in obesity
Abstract
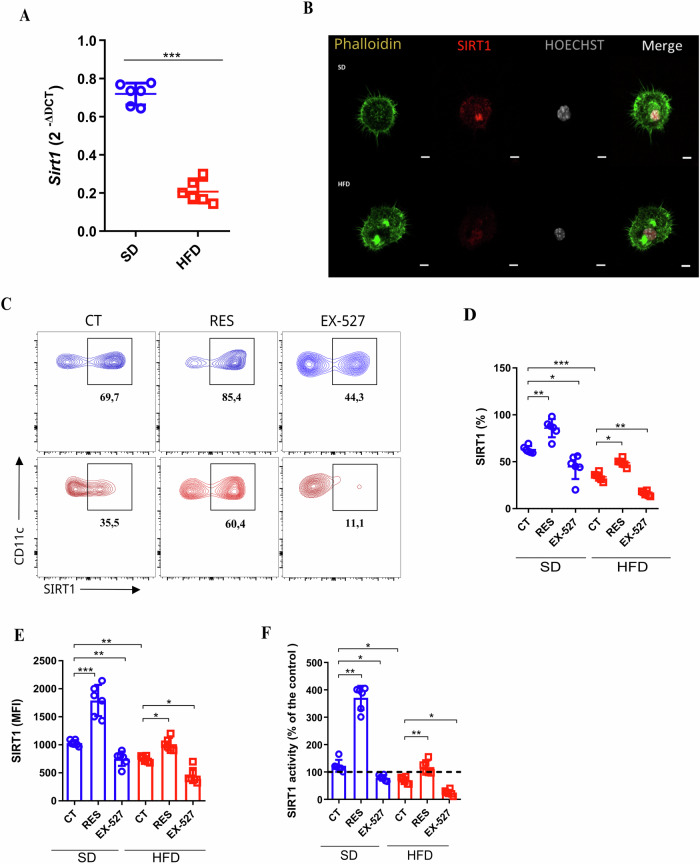
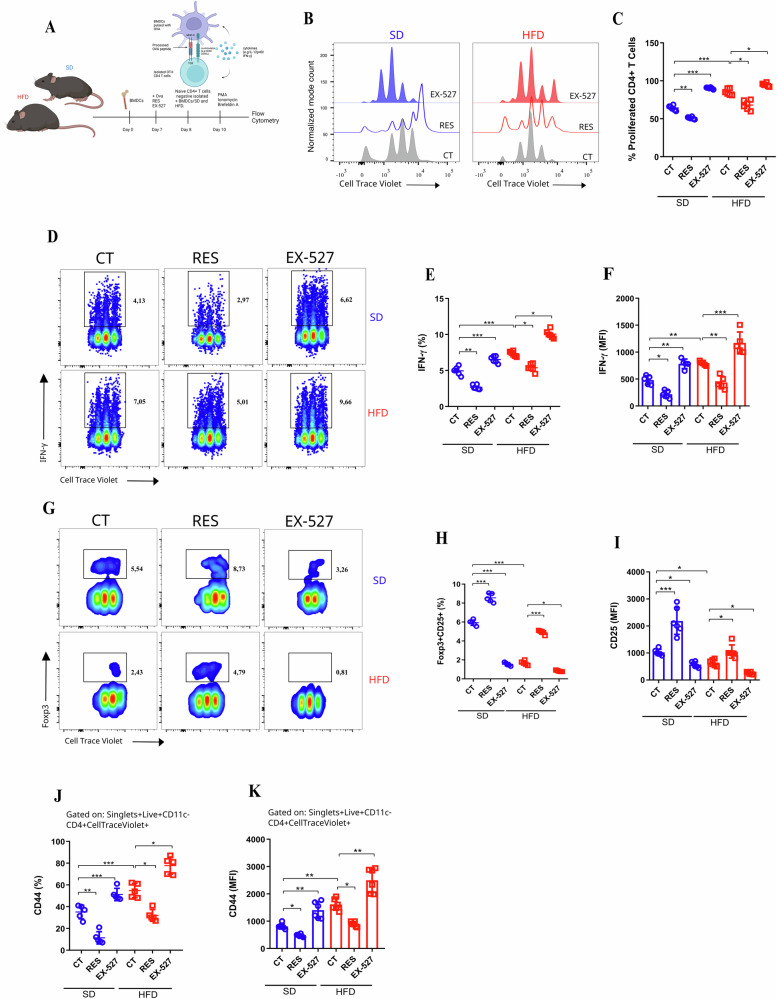
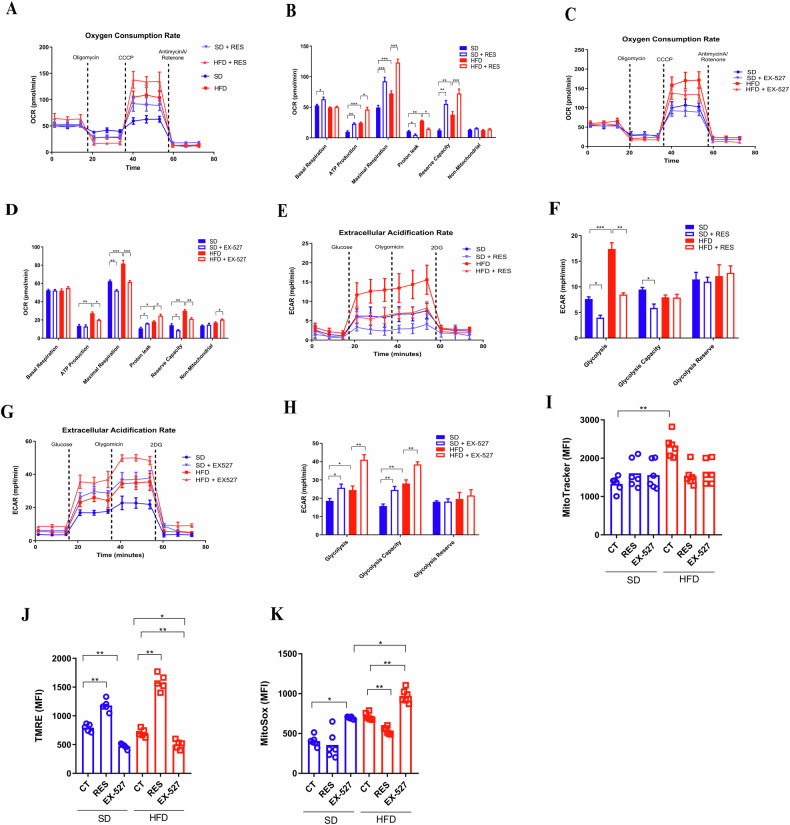
Sirtuin 1 (SIRT1) is a class III histone deacetylase (HDAC3) that plays a crucial role in regulating the activation and differentiation of dendritic cells (DCs) as well as controlling the polarization and activation of T cells. Obesity, a chronic inflammatory condition, is characterized by the activation of immune cells in various tissues. We hypothesized that SIRT1 might influence the phenotype and functions of DCs through the Ido1 pathway, ultimately leading to the polarization towards pro-inflammatory T cells in obesity. In our study, we observed that SIRT1 activity was reduced in bone marrow-derived DCs (BMDCs) from obese animals. These BMDCs exhibited elevated oxidative phosphorylation (OXPHOS) and increased extracellular acidification rates (ECAR), along with enhanced expression of class II MHC, CD86, and CD40, and elevated secretion of IL-12p40, while the production of TGF-β was reduced. The kynurenine pathway activity was decreased in BMDCs from obese animals, particularly under SIRT1 inhibition. SIRT1 positively regulated the expression of Ido1 in DCs in a PPARγ-dependent manner. To support these findings, ATAC-seq analysis revealed that BMDCs from obese mice had differentially regulated open chromatin regions compared to those from lean mice, with reduced chromatin accessibility at the Sirt1 genomic locus in BMDCs from obese WT mice. Gene Ontology (GO) enrichment analysis indicated that BMDCs from obese animals had disrupted metabolic pathways, including those related to GTPase activity and insulin response. Differential expression analysis showed reduced levels of Pparg and Sirt1 in BMDCs from obese mice, which was challenged and confirmed using BMDCs from mice with conditional knockout of Sirt1 in dendritic cells (SIRT1∆). This study highlights that SIRT1 controls the metabolism and functions of DCs through modulation of the kynurenine pathway, with significant implications for obesity-related inflammation.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Asfaha Y, Schrenk C, Alves Avelar LA, Hamacher A, Pflieger M, Kassack MU, et al. Recent advances in class IIa histone deacetylases research. Bioorg Med Chem. 2019;27:115087. - PubMed
-
- Leite JA, Ghirotto B, Targhetta VP, de Lima J, Câmara NOS. Sirtuins as pharmacological targets in neurodegenerative and neuropsychiatric disorders. Br J Pharmacol. 2022;179:1496–511. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
