

Integrated stress responses in okra plants (cv. ''Meya']: unravelling the mechanisms underlying drought and nematode co-occurrence
- PMID: 39427110
- PMCID: PMC11490165
- DOI: 10.1186/s12870-024-05686-1
Integrated stress responses in okra plants (cv. ''Meya']: unravelling the mechanisms underlying drought and nematode co-occurrence
Erratum in
-
Correction: Integrated stress responses in okra plants (cv. ''Meya']: unravelling the mechanisms underlying drought and nematode cooccurrence.BMC Plant Biol. 2024 Oct 29;24(1):1021. doi: 10.1186/s12870-024-05750-w. BMC Plant Biol. 2024. PMID: 39468447 Free PMC article. No abstract available.
Abstract
Background: Climate change threatens sub-Saharan Africa's agricultural production, causing abiotic and biotic stressors. The study of plant responses to joint stressors is crucial for understanding molecular processes and identifying resilient crops for global food security. This study aimed to explore the shared and tailored responses of okra plants (cv. ''Meya'), at the biochemical and molecular levels, subjected to combined stresses of drought and Meloidogyne incognita infection.
Design: The study involved 240 okra plants in a completely randomized design, with six treatments replicated 20 times. Okra plants were adequately irrigated at the end of every 10-days water deficit that lasted for 66 days (D). Also, the plants were infected with M. incognita for 66 days and irrigated at 2-days intervals (R). The stresses were done independently, in sequential combination (D before R and R before D) and concurrently (R and D). All biochemical and antioxidant enzyme assays were carried out following standard procedures.
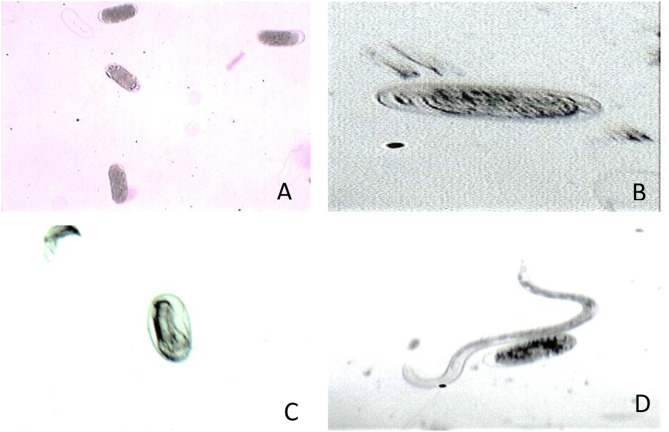
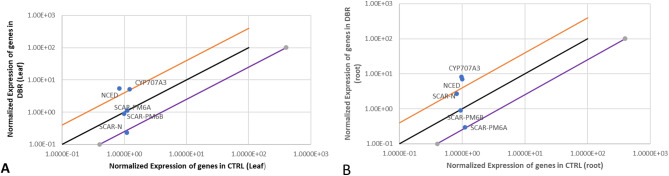
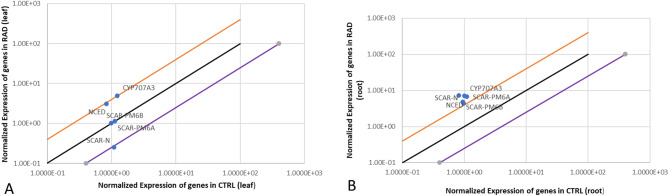
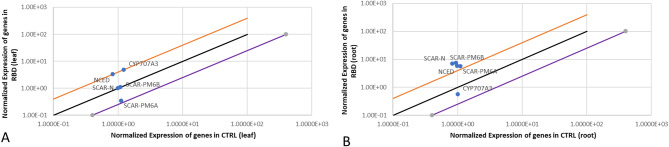
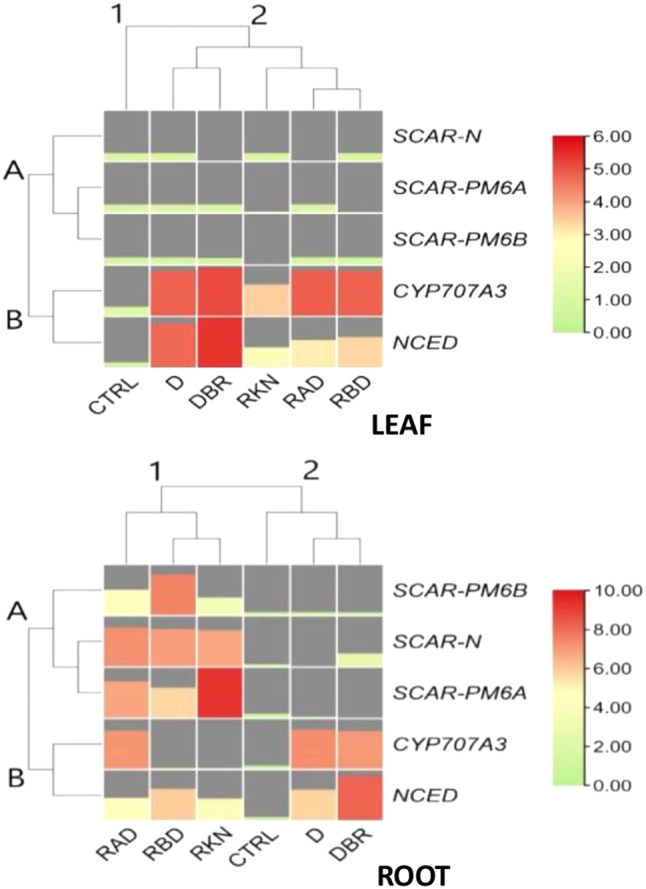
Results: Significant reductions in leaf relative water content were recorded in all stressed plants, especially in leaves of plants under individual drought stress (D) (41.6%) and plants stressed with root-knot nematode infection before drought stress (RBD) (41.4%). Malondialdehyde contents in leaf tissues from plants in D, nematode-only stress (RKN), drought stress before root-knot nematode infection (DBR), RBD, and concurrent drought-nematode stress (RAD) significantly increased by 320.2%, 152.9%, 186.5%, 283.7%, and 109.6%, respectively. Plants in D exhibited the highest superoxide dismutase activities in leaf (147.1% increase) and root (105.8% increase) tissues. Catalase (CAT) activities were significantly increased only in leaves of plants in D (90.8%) and RBD (88.9%), while only roots of plants in D exhibited a substantially higher CAT activity (139.3% increase) in comparison to controlled plants. Okra plants over-expressed NCED3 and under-expressed Me3 genes in leaf tissues. The NCED3 gene was overexpressed in roots from all treatments, while CYP707A3 was under-expressed only in roots of plants in RBD and RKN. CYP707A3 and NCED3 were grouped as closely related genes, while members of the Me3 genes were clustered into a separate group.
Conclusion: The biochemical and molecular responses observed in okra plants (cv. ''Meya') subjected to combined stresses of drought and Meloidogyne incognita infection provide valuable insights into enhancing crop resilience under multifaceted stress conditions, particularly relevant for agricultural practices in sub-Saharan Africa facing increasing climatic challenges.
Keywords: Meloidogyne incognita; Biotic and abiotic stress; Drought; Gene regulation; Okra.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Anzano A, Bonanomi G, Mazzoleni S, Lanzotti V. Plant metabolomics in biotic and abiotic stress: a critical overview. Phytochem Rev. 2021;21:503–24. 10.1007/s11101-021-09786-w.
-
- Santos TBD, Ribas AF, De Souza SGH, Budzinski IGF, Domingues DS. Physiological responses to drought, salinity, and heat stress in plants: a review. Stresses. 2022;2:113–35. 10.3390/stresses2010009.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
