

Conformational cycle of a protease-containing ABC transporter in lipid nanodiscs reveals the mechanism of cargo-protein coupling
- PMID: 39428489
- PMCID: PMC11491471
- DOI: 10.1038/s41467-024-53420-0
Conformational cycle of a protease-containing ABC transporter in lipid nanodiscs reveals the mechanism of cargo-protein coupling
Abstract
Protease-containing ABC transporters (PCATs) couple the energy of ATP hydrolysis to the processing and export of diverse cargo proteins across cell membranes to mediate antimicrobial resistance and quorum sensing. Here, we combine biochemical analysis, single particle cryoEM, and DEER spectroscopy in lipid bilayers along with computational analysis to illuminate the structural and energetic underpinnings of coupled cargo protein export. Our integrated investigation uncovers competitive interplay between nucleotides and cargo protein binding that ensures the latter's orderly processing and subsequent transport. The energetics of cryoEM structures in lipid bilayers are congruent with the inferred mechanism from ATP turnover analysis and reveal a snapshot of a high-energy outward-facing conformation that provides an exit pathway into the lipid bilayer and/or the extracellular side. DEER investigation of the core ABC transporter suggests evolutionary tuning of the energetic landscape to fulfill the function of substrate processing prior to export.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

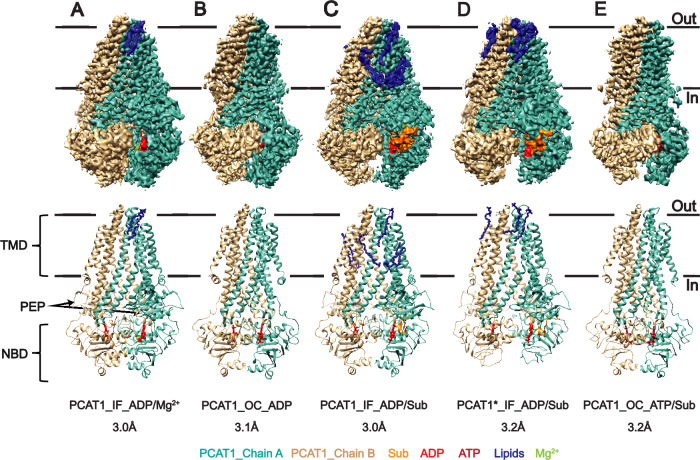
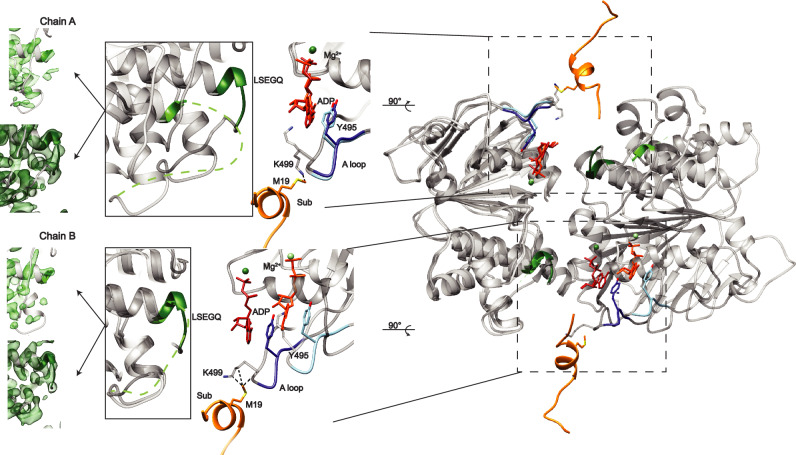
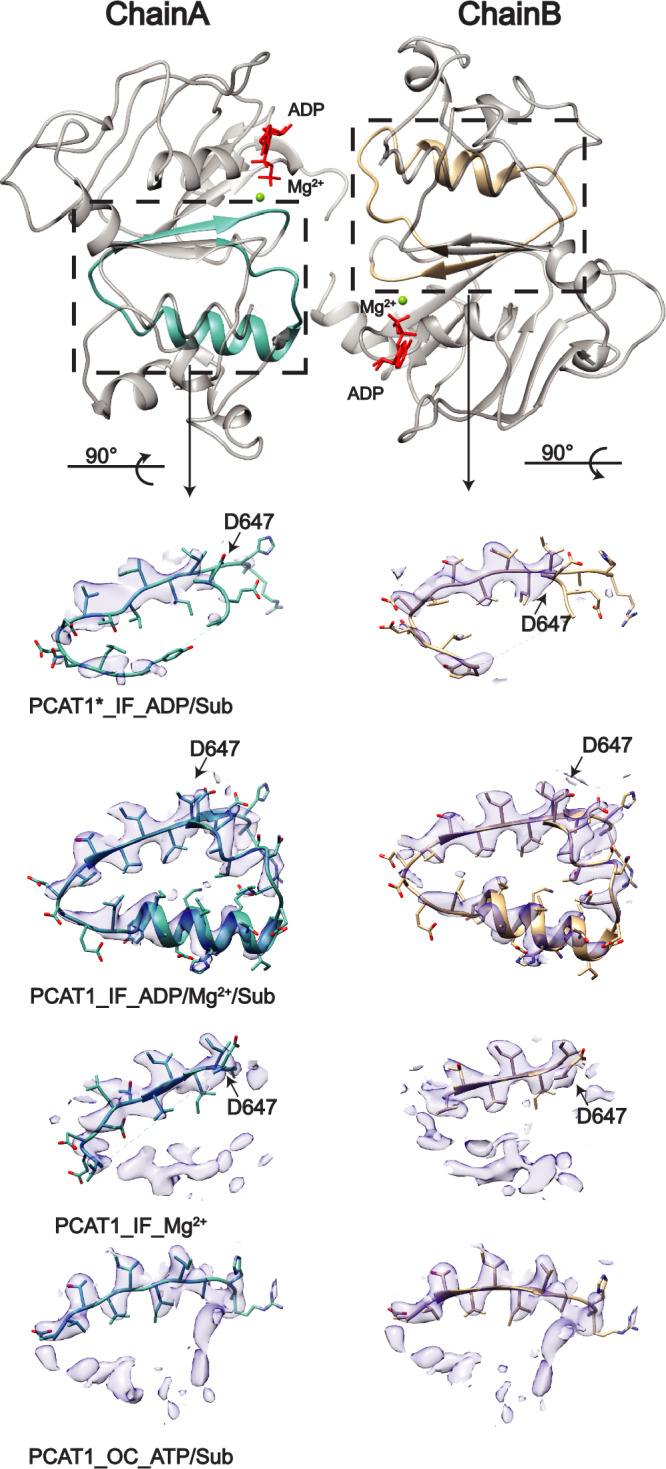
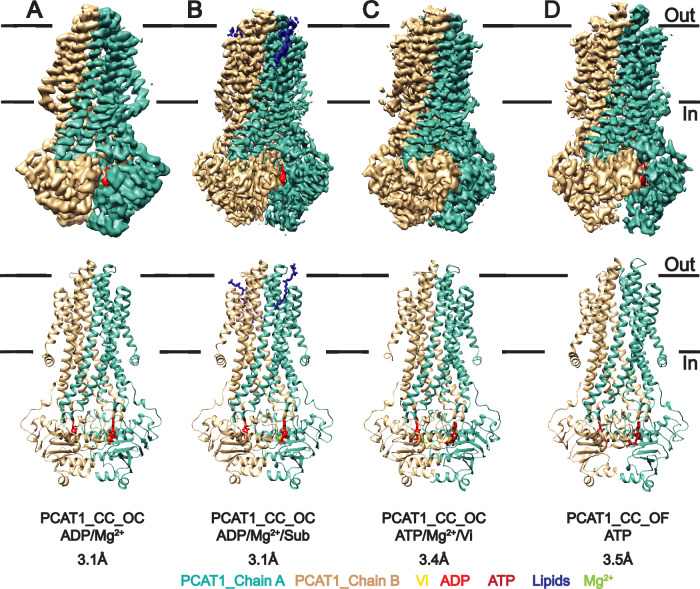
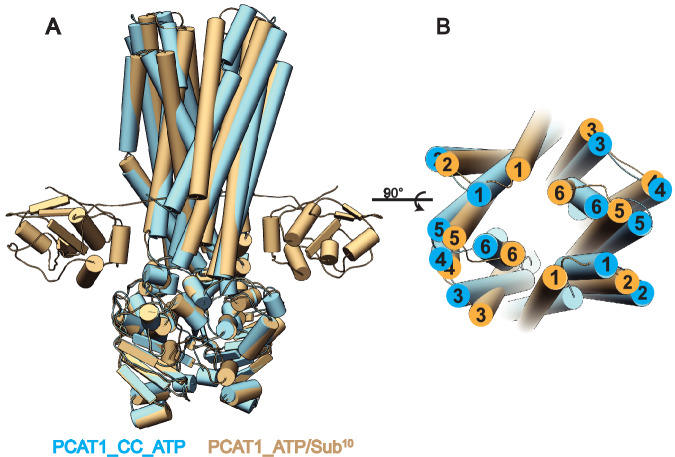
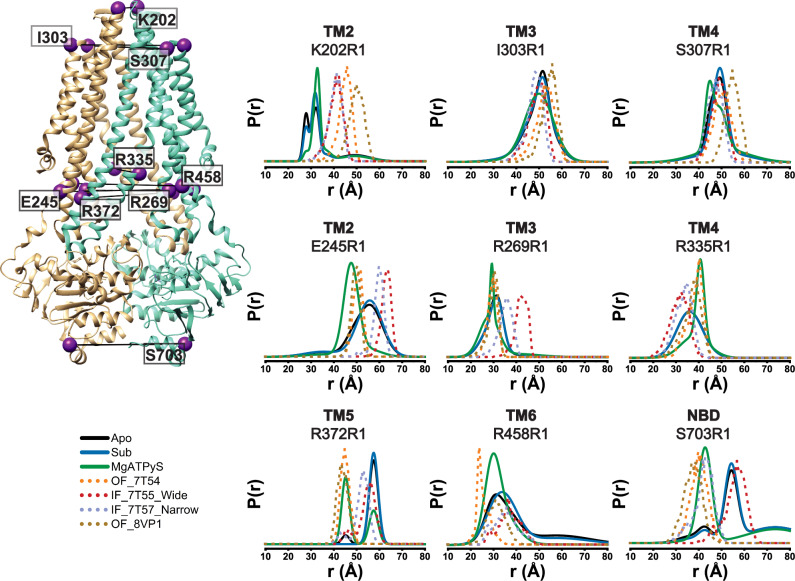
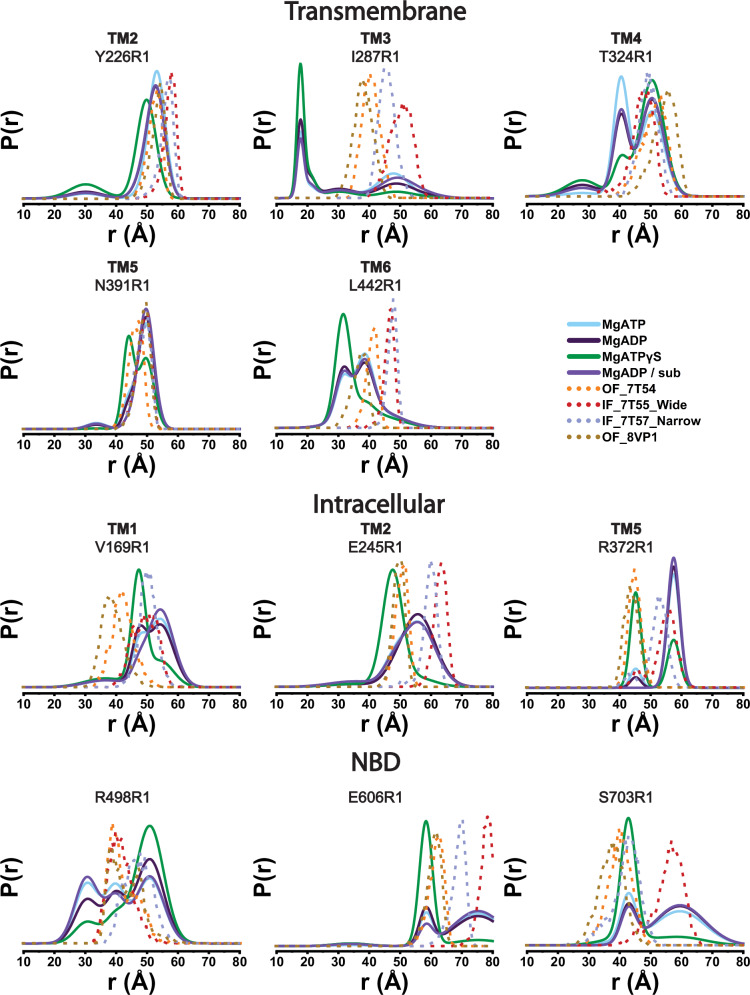
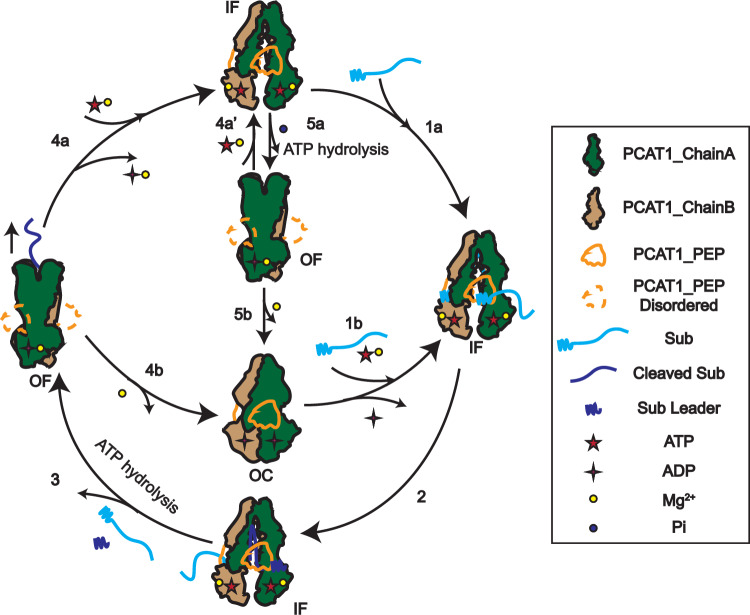
References
-
- Thomas, C. & Tampe, R. Structural and mechanistic principles of ABC transporters. Annu Rev. Biochem.89, 605–636 (2020). - PubMed
-
- Locher, K. P. Mechanistic diversity in ATP-binding cassette (ABC) transporters. Nat. Struct. Mol. Biol.23, 487–493 (2016). - PubMed
-
- Gebhard, S. A. B. C. transporters of antimicrobial peptides in Firmicutes bacteria - phylogeny, function and regulation. Mol. Microbiol.86, 1295–1317 (2012). - PubMed
-
- Gebhard, S. & Mascher, T. Antimicrobial peptide sensing and detoxification modules: unravelling the regulatory circuitry of Staphylococcus aureus. Mol. Microbiol.81, 581–587 (2011). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
