

SUMOylation regulates the aggressiveness of breast cancer-associated fibroblasts
- PMID: 39432155
- PMCID: PMC11996949
- DOI: 10.1007/s13402-024-01005-w
SUMOylation regulates the aggressiveness of breast cancer-associated fibroblasts
Abstract
Background: Cancer-associated fibroblasts (CAFs) are the most abundant stromal cellular component in the tumor microenvironment (TME). CAFs contribute to tumorigenesis and have been proposed as targets for anticancer therapies. Similarly, dysregulation of SUMO machinery components can disrupt the balance of SUMOylation, contributing to tumorigenesis and drug resistance in various cancers, including breast cancer. We explored the role of SUMOylation in breast CAFs and evaluated its potential as a therapeutic strategy in breast cancer.
Methods: We used pharmacological and genetic approaches to analyse the functional crosstalk between breast tumor cells and CAFs. We treated breast CAFs with the SUMO1 inhibitor ginkgolic acid (GA) at two different concentrations and conditioned media was used to analyse the proliferation, migration, and invasion of breast cancer cells from different molecular subtypes. Additionally, we performed quantitative proteomics (SILAC) to study the differential signalling pathways expressed in CAFs treated with low or high concentrations of GA. We confirmed these results both in vitro and in vivo. Moreover, we used samples from metastatic breast cancer patients to evaluate the use of GA as a therapeutic strategy.
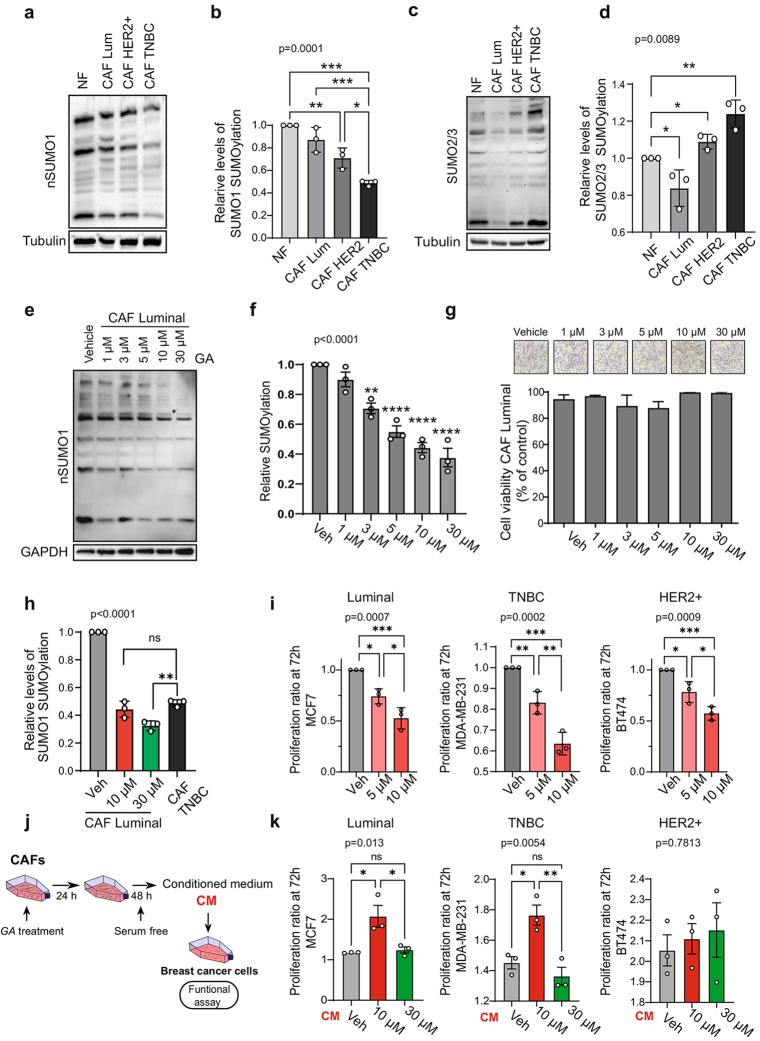
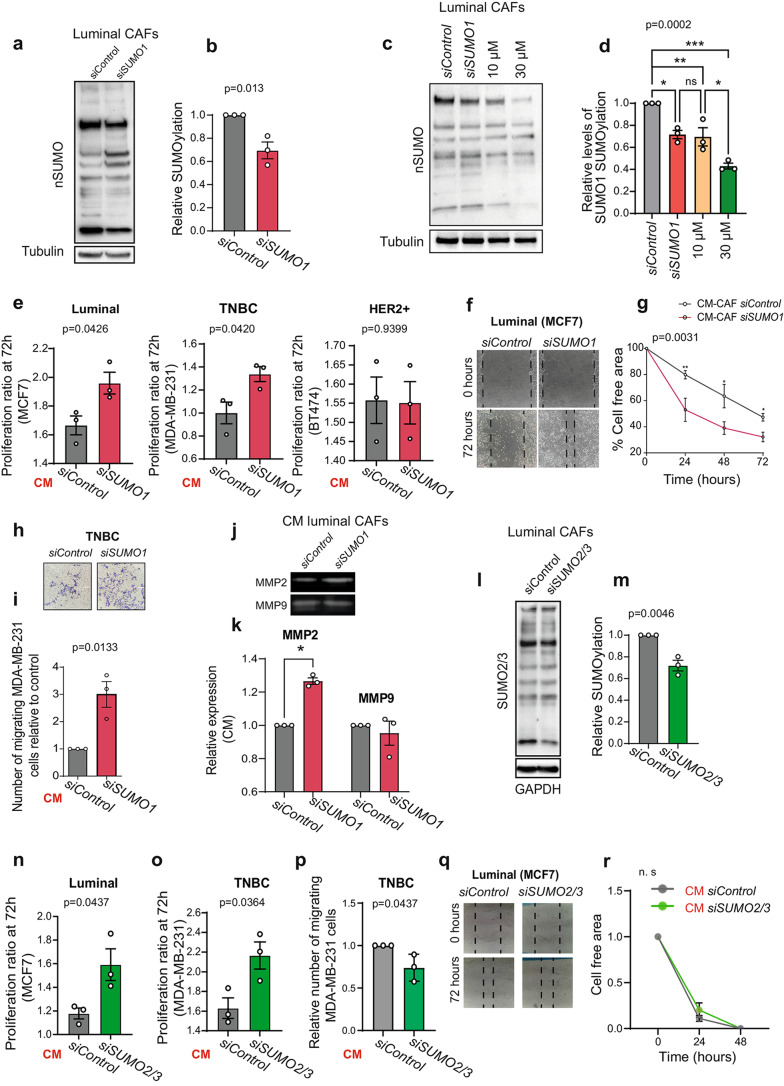
Results: Inhibition of SUMOylation with ginkgolic acid (GA) induces death in breast cancer cells but does not affect the viability of CAFs, indicating that CAFs are resistant to this therapy. While CAF viability is unaffected, CAF-conditioned media (CM) is altered by GA, impacting tumor cell behaviour in different ways depending on the overall degree to which SUMO1-SUMOylated proteins are dysregulated. Breast cancer cell lines exhibited a concentration-dependent response to conditioned media (CM) from CAFs. At a low concentration of GA (10 µM), there was an increase in proliferation, migration and invasion of breast cancer cells. However, at a higher concentration of GA (30 µM), these processes were inhibited. Similarly, analysis of tumor development revealed that at 10 µM of GA, the tumors were heavier and there was a greater degree of metastasis compared to the tumors treated with the higher concentration of GA (30 µM). Moreover, some of these effects could be explained by an alteration in the activity of the GTPase Rac1 and the activation of the AKT signalling pathway. The results obtained using SILAC suggest that different concentrations of GA affected cellular processes differentially, possibly influencing the secretome of CAFs. Treatment of metastatic breast cancer with GA demonstrated the use of SUMOylation inhibition as an alternative therapeutic strategy.
Conclusion: The study highlights the importance of SUMOylation in the tumor microenvironment, specifically in cancer-associated fibroblasts (CAFs). Targeting SUMOylation in CAFs affects their signalling pathways and secretome in a concentration-dependent manner, regulating the protumorigenic properties of CAFs.
Keywords: Breast cancer; CAFs; Ginkgolic acid; SILAC; SUMOylation.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Ascites effusions were collected by the Manchester Cancer Research Centre (MCRC) Biobank (ethical approval: 12-ROCL-01). Written informed consent was obtained from each patient. Competing interests: The authors declare no competing interests.
Figures




References
-
- V. Bernier-Villamor, et al., Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell 108(3), 345–356 (2002) - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
