

Comparing neoantigen cancer vaccines and immune checkpoint therapy unveils an effective vaccine and anti-TREM2 macrophage-targeting dual therapy
- PMID: 39446585
- PMCID: PMC11785356
- DOI: 10.1016/j.celrep.2024.114875
Comparing neoantigen cancer vaccines and immune checkpoint therapy unveils an effective vaccine and anti-TREM2 macrophage-targeting dual therapy
Abstract
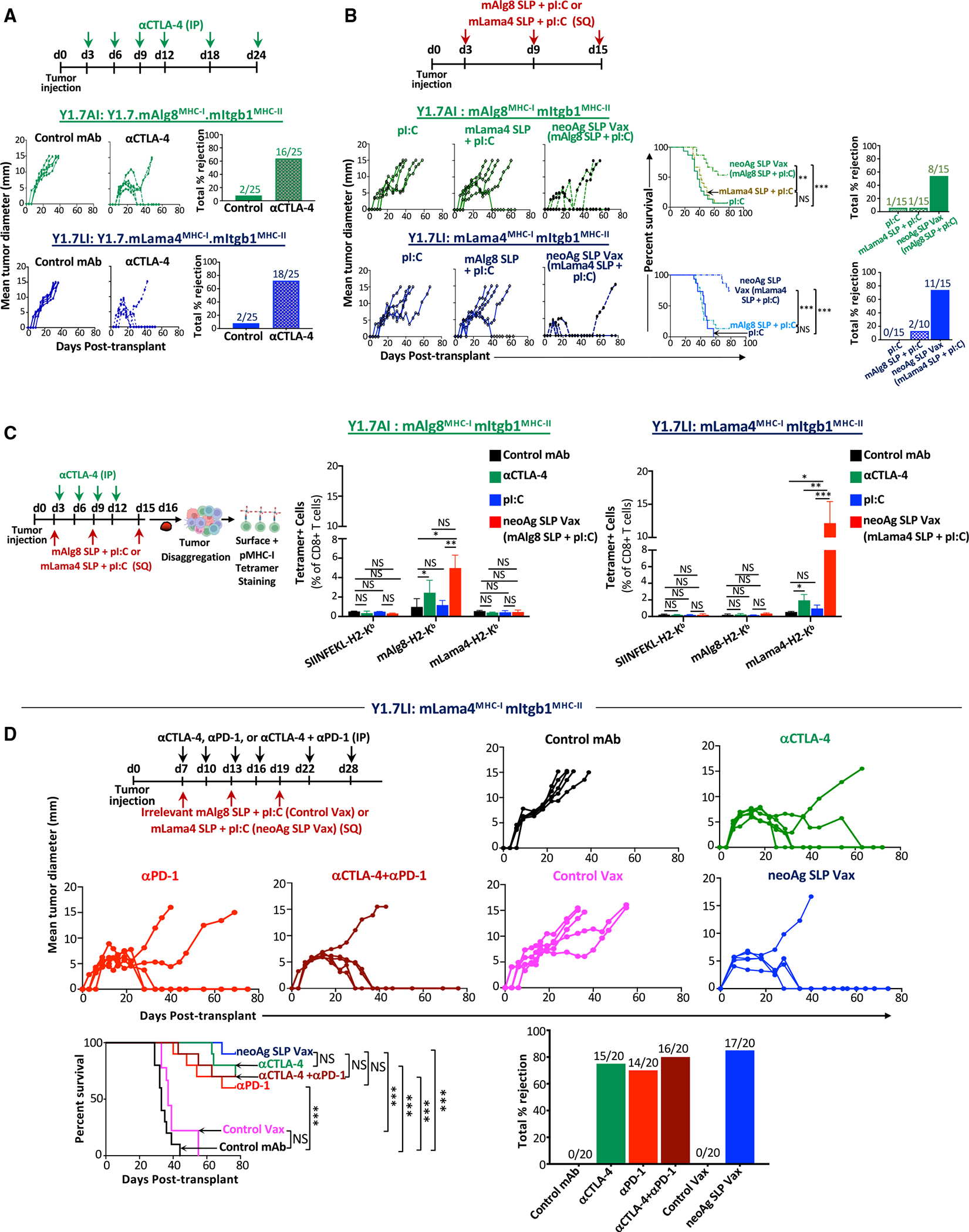
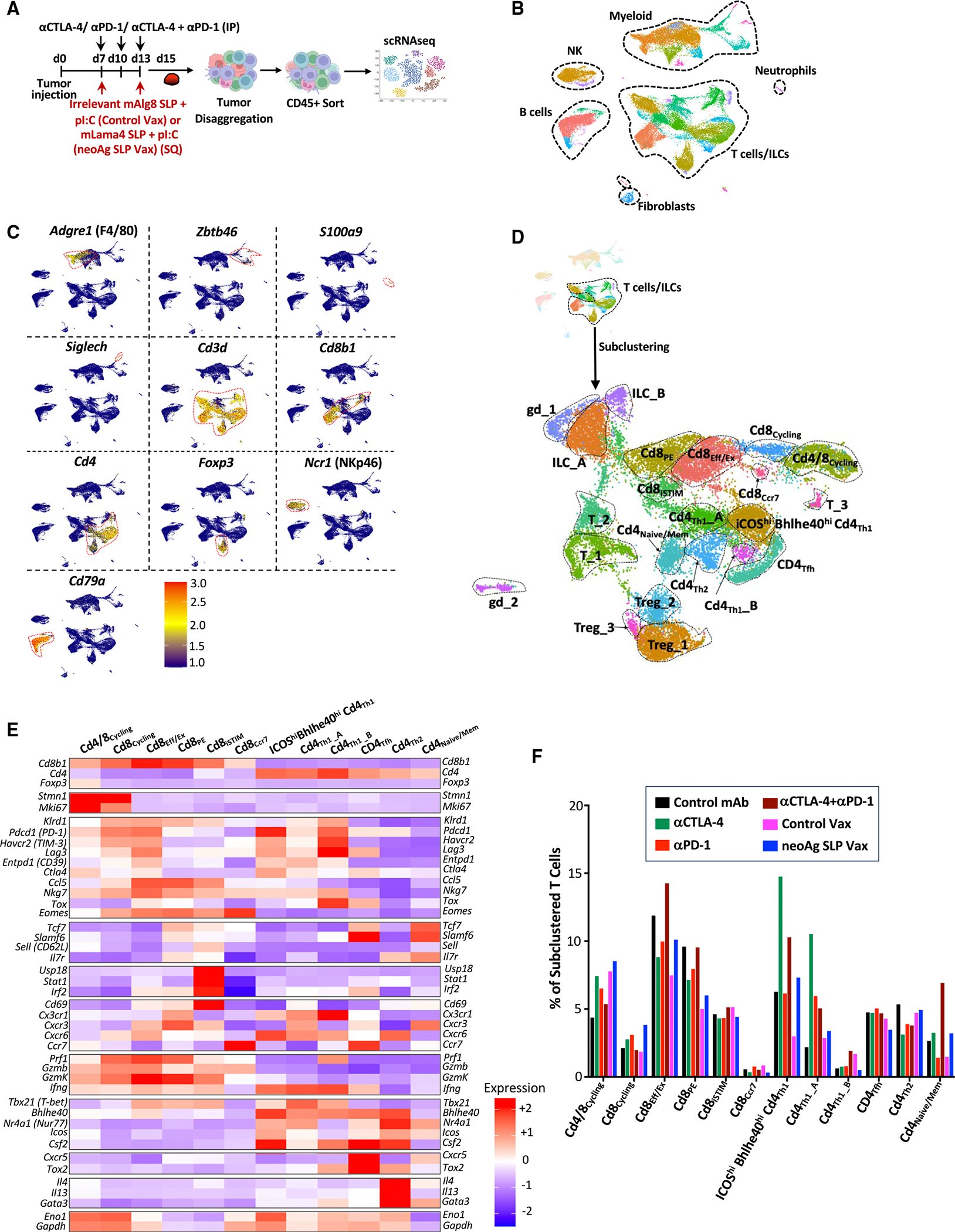
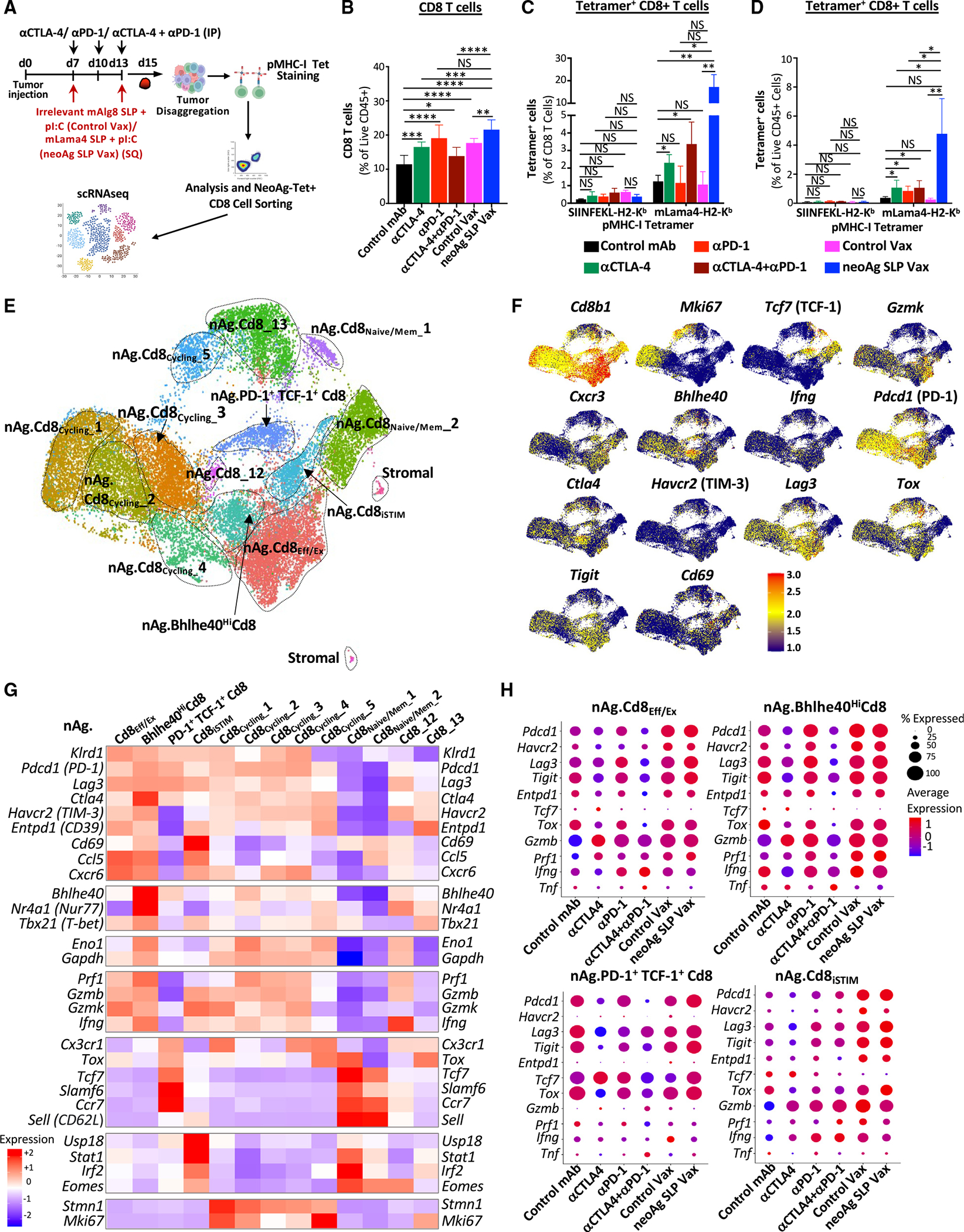
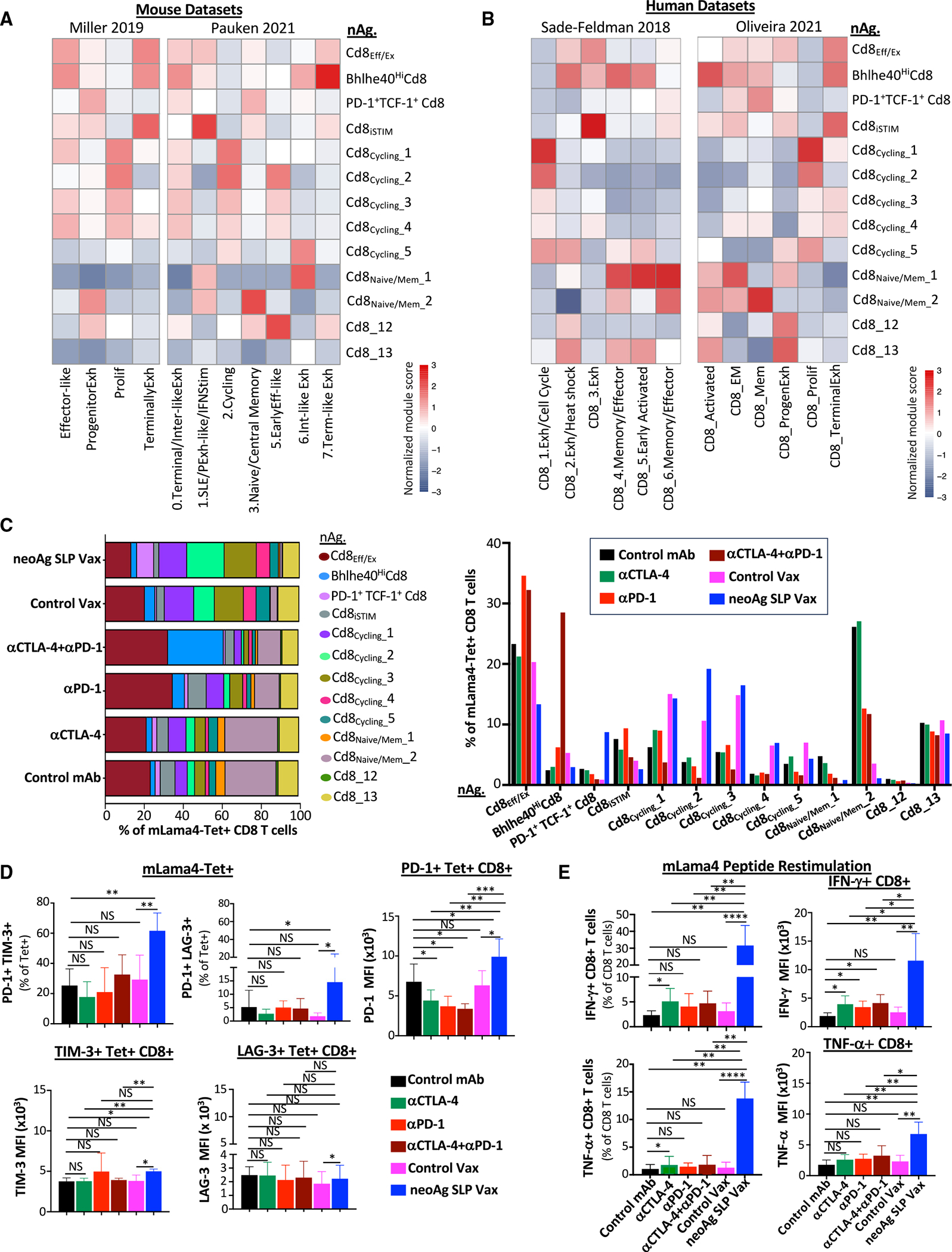
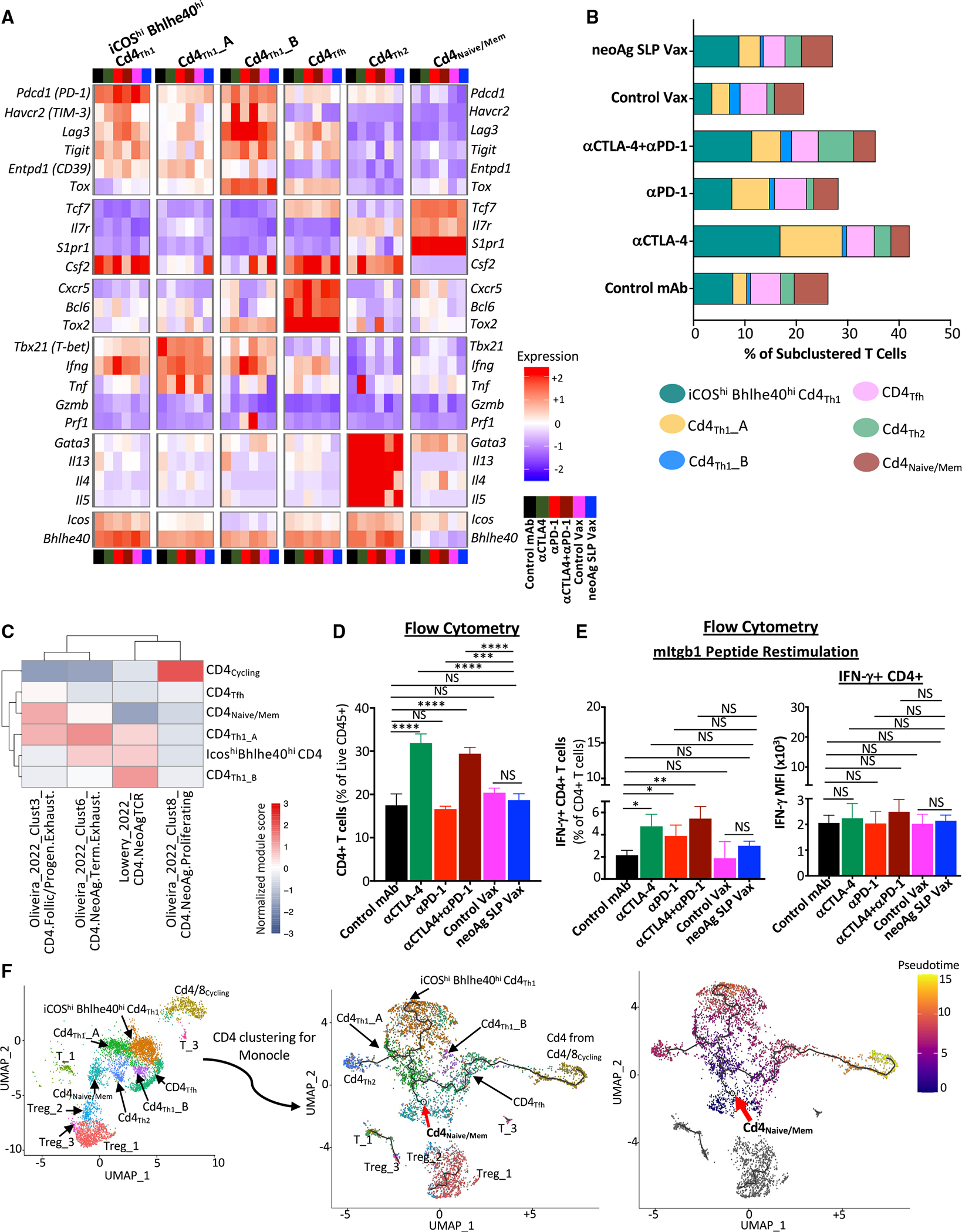
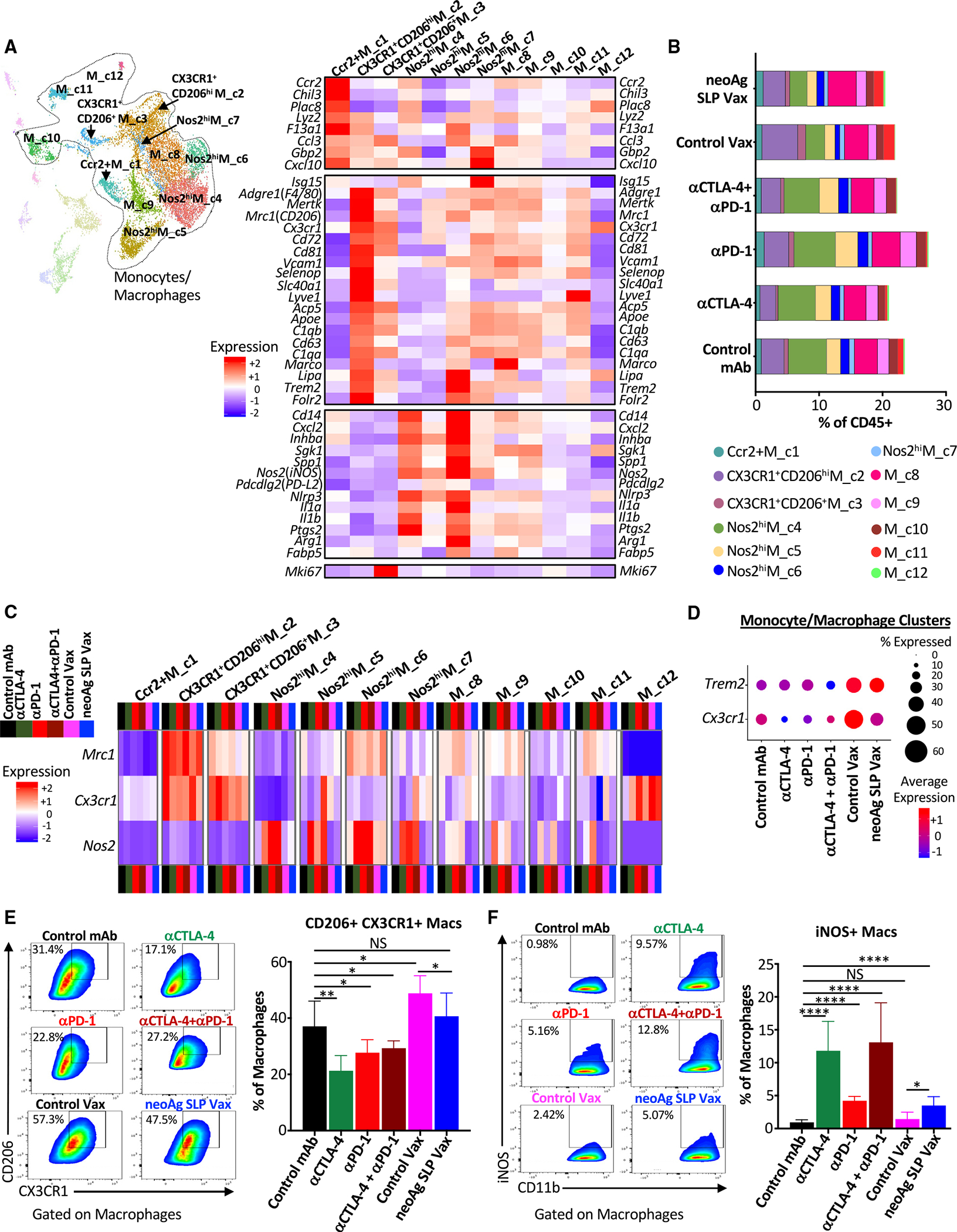
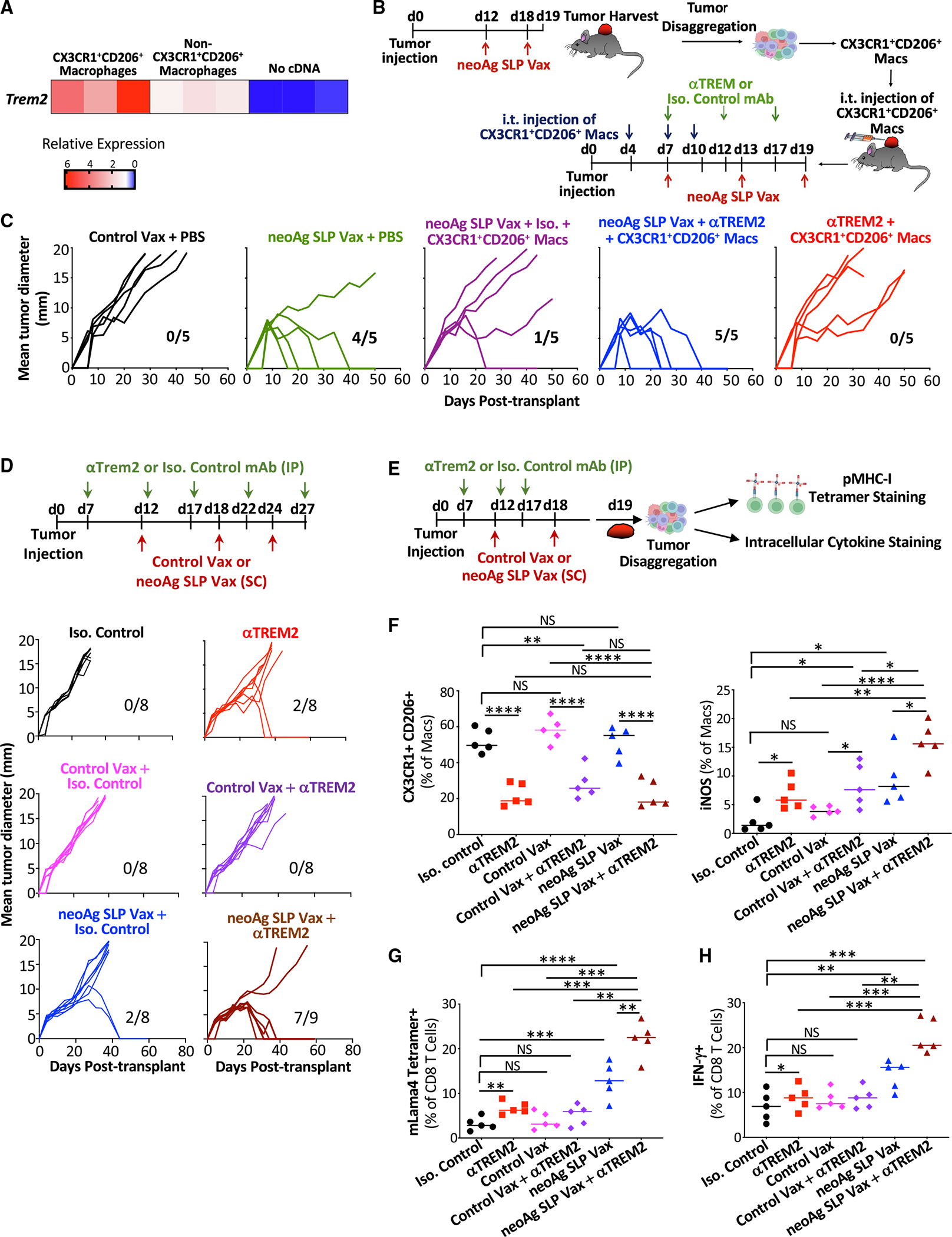
The goal of therapeutic cancer vaccines and immune checkpoint therapy (ICT) is to promote T cells with anti-tumor capabilities. Here, we compared mutant neoantigen (neoAg) peptide-based vaccines with ICT in preclinical models. NeoAg vaccines induce the most robust expansion of proliferating and stem-like PD-1+TCF-1+ neoAg-specific CD8 T cells in tumors. Anti-CTLA-4 and/or anti-PD-1 ICT promotes intratumoral TCF-1- neoAg-specific CD8 T cells, although their phenotype depends in part on the specific ICT used. Anti-CTLA-4 also prompts substantial changes to CD4 T cells, including induction of ICOS+Bhlhe40+ T helper 1 (Th1)-like cells. Although neoAg vaccines or ICTs expand iNOS+ macrophages, neoAg vaccines maintain CX3CR1+CD206+ macrophages expressing the TREM2 receptor, unlike ICT, which suppresses them. TREM2 blockade enhances neoAg vaccine efficacy and is associated with fewer CX3CR1+CD206+ macrophages and induction of neoAg-specific CD8 T cells. Our findings highlight different mechanisms underlying neoAg vaccines and different forms of ICT and identify combinatorial therapies to enhance neoAg vaccine efficacy.
Keywords: CD4 T cells; CP: Cancer; CP: Immunology; TREM2; anti-CTLA-4/anti-PD-1; cancer immunotherapy; combination immunotherapy; immune checkpoint therapy; intratumoral macrophages; neoantigen cancer vaccines; neoantigen-specific CD8 T cells; tumor microenvironment.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.M.H. is currently employed by Lifecycle Biotechnologies, and her employment does not pose any direct conflict of interest with the research presented in this article. M.M.G. reports a personal honorarium of US $1,000.00 per year from Springer Nature Ltd (as an associate editor for Nature Precision Oncology) and served as a paid consultant for Merck. M. Colonna reports that they are a member of the scientific advisory board of Vigil Neuro and Cell Signaling Technology, received research grants from Vigil Neuro during the conduct of the study, and has a patent related to TREM2 modulation pending (PCT/US2021/019914). K.E.P. reports an advising relationship with Guardant Health that will result in advising fees.
Figures
Update of
-
Neoantigen Cancer Vaccines and Different Immune Checkpoint Therapies Each Utilize Both Converging and Distinct Mechanisms that in Combination Enable Synergistic Therapeutic Efficacy.bioRxiv [Preprint]. 2024 Jan 26:2023.12.20.570816. doi: 10.1101/2023.12.20.570816. bioRxiv. 2024. Update in: Cell Rep. 2024 Nov 26;43(11):114875. doi: 10.1016/j.celrep.2024.114875. PMID: 38187708 Free PMC article. Updated. Preprint.
References
-
- Carreno BM, Magrini V, Becker-Hapak M, Kaabinejadian S, Hundal J, Petti AA, Ly A, Lie WR, Hildebrand WH, Mardis ER, and Linette GP (2015). A dendritic cell vaccine increases the breadth and diversity of melanoma neoantigen-specific T cells. Science 348, 803–808. 10.1126/science.aaa3828. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
