

Retrotransposons are co-opted to activate hematopoietic stem cells and erythropoiesis
- PMID: 39446896
- PMCID: PMC11709122
- DOI: 10.1126/science.ado6836
Retrotransposons are co-opted to activate hematopoietic stem cells and erythropoiesis
Abstract
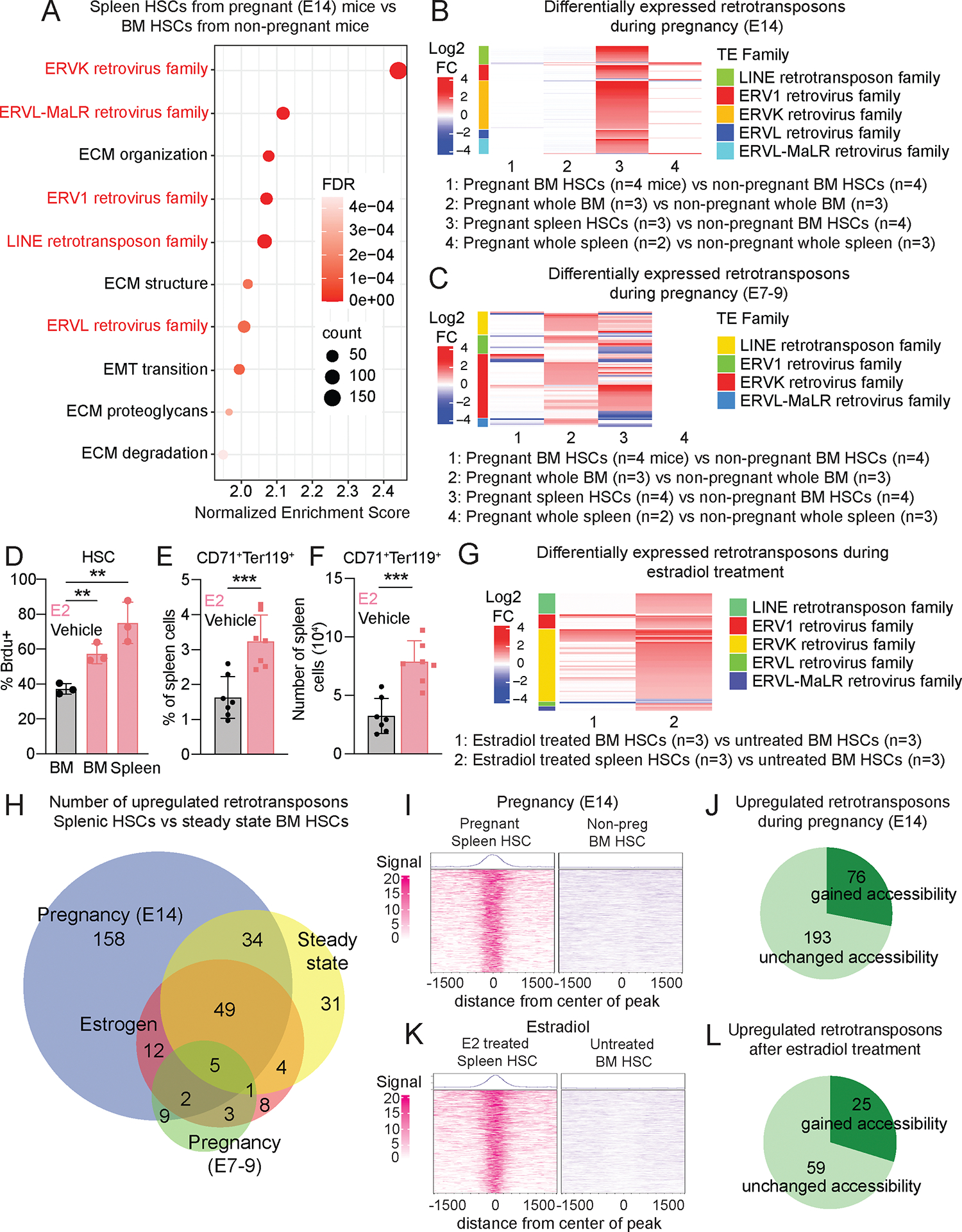
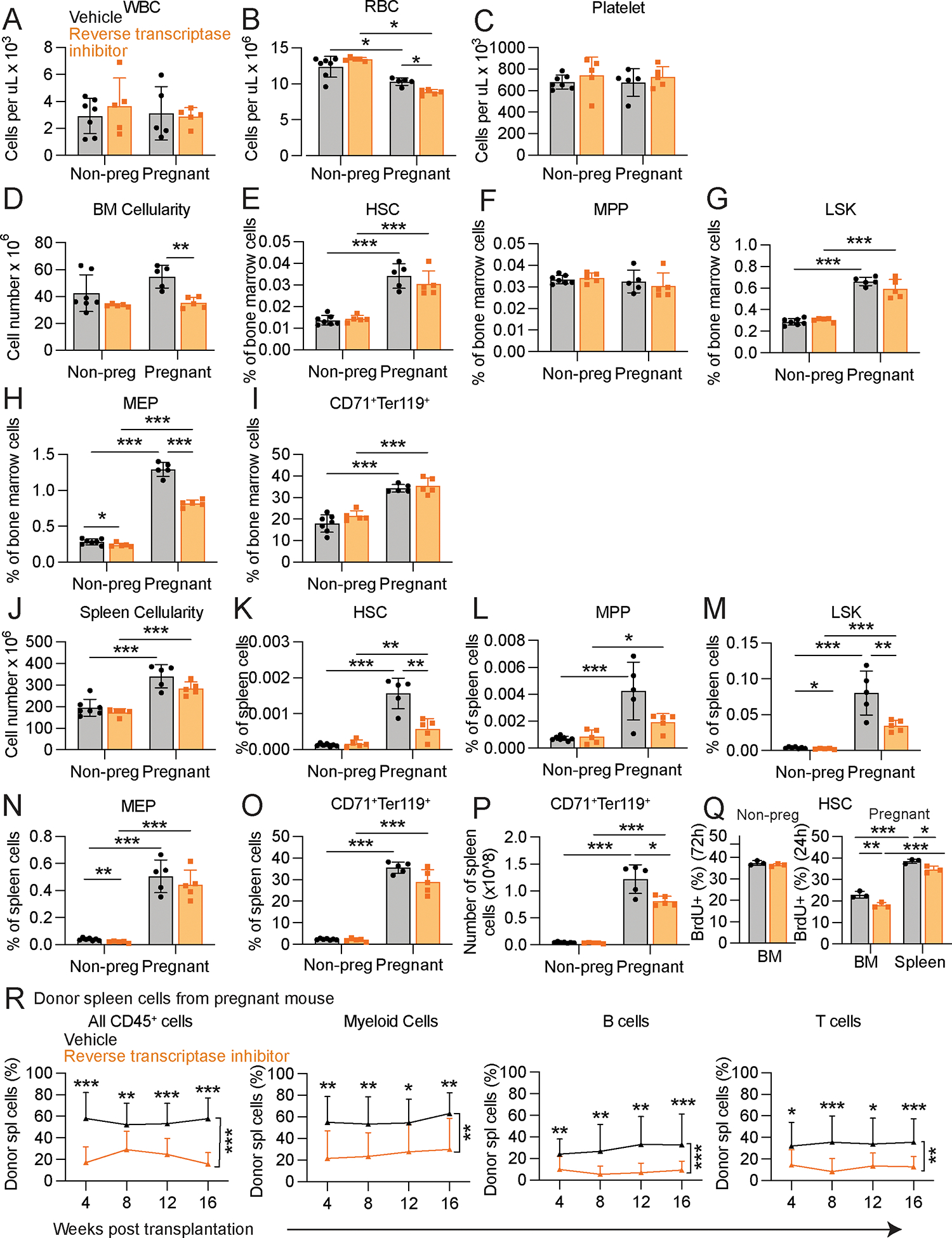
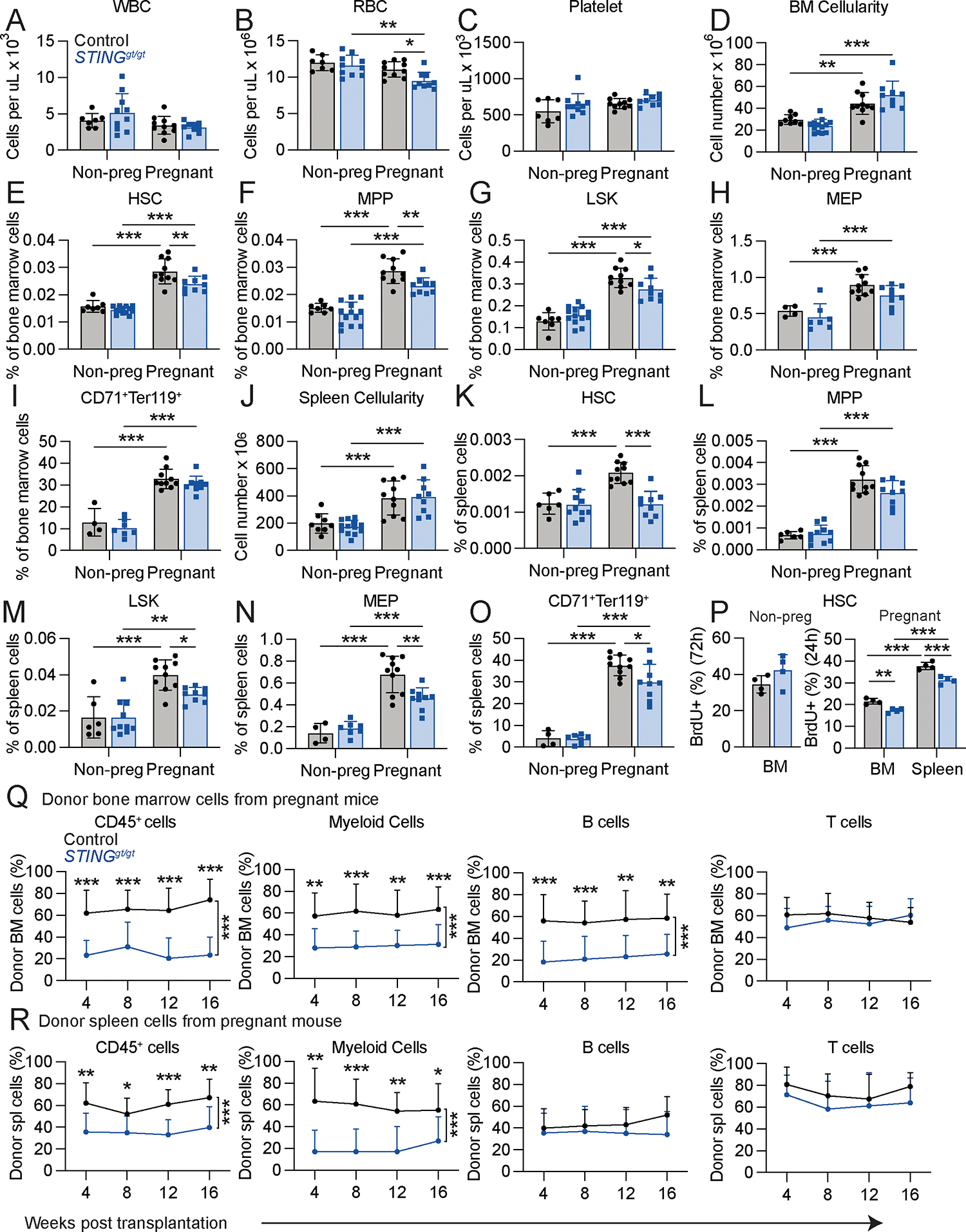
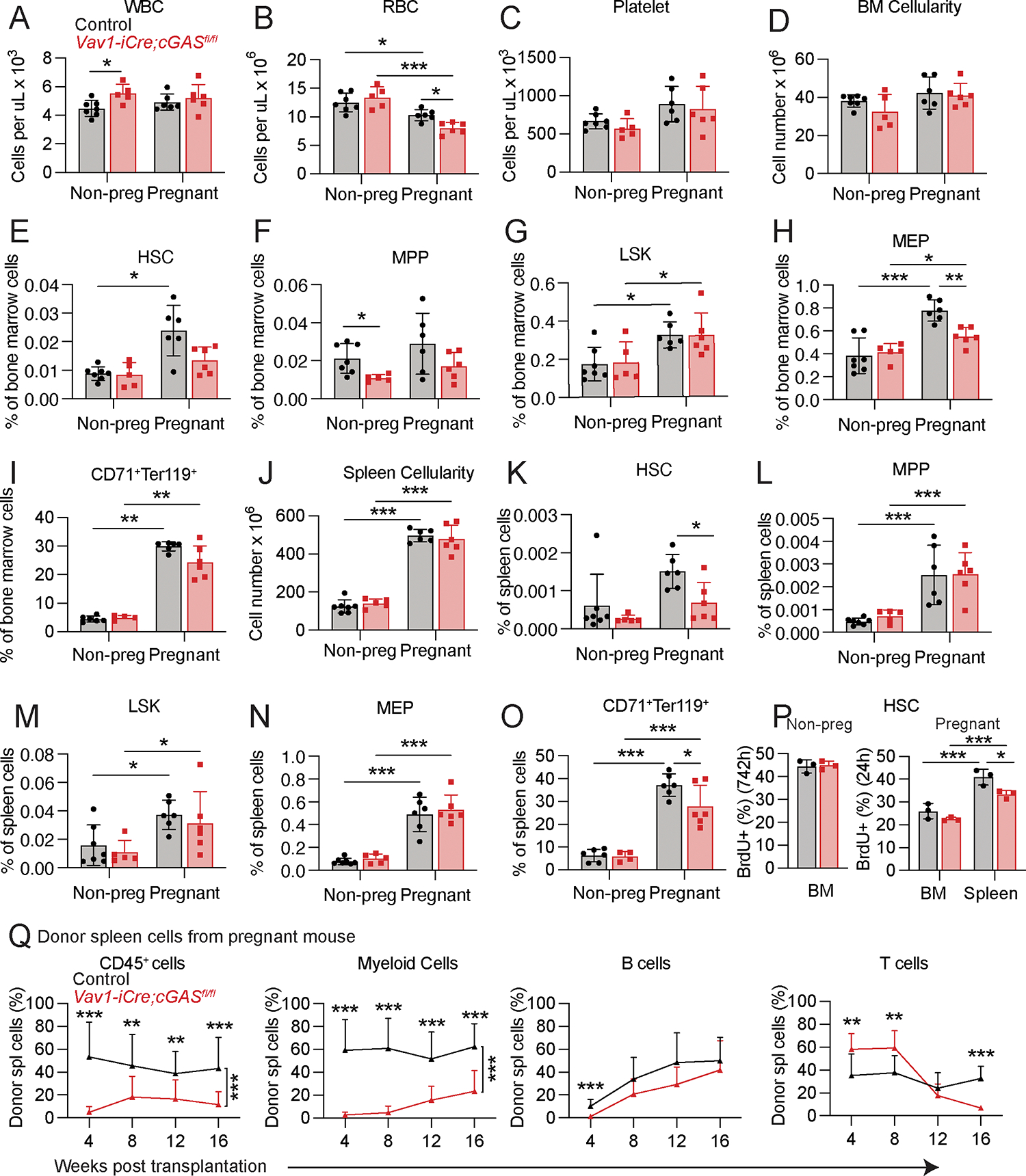
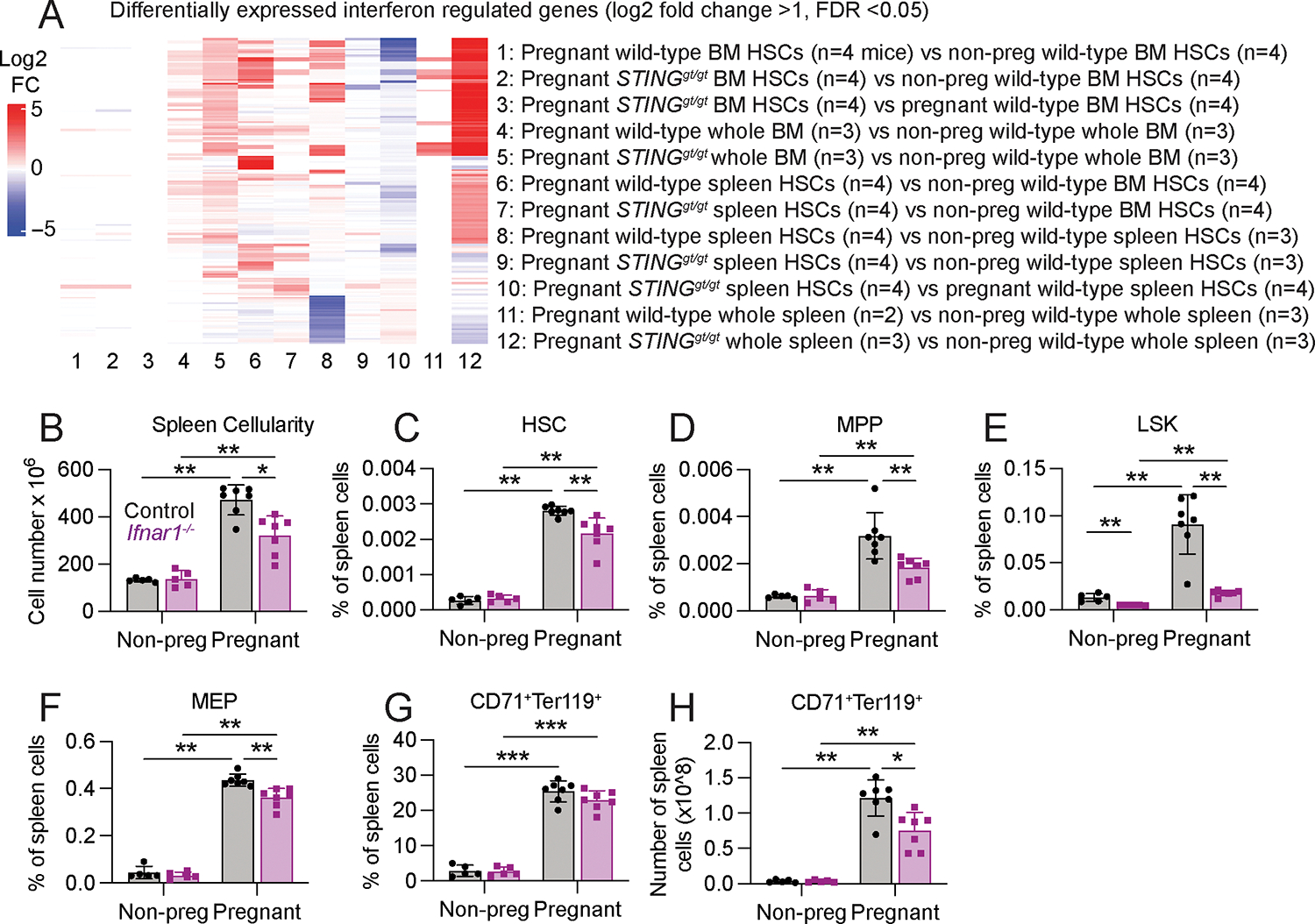
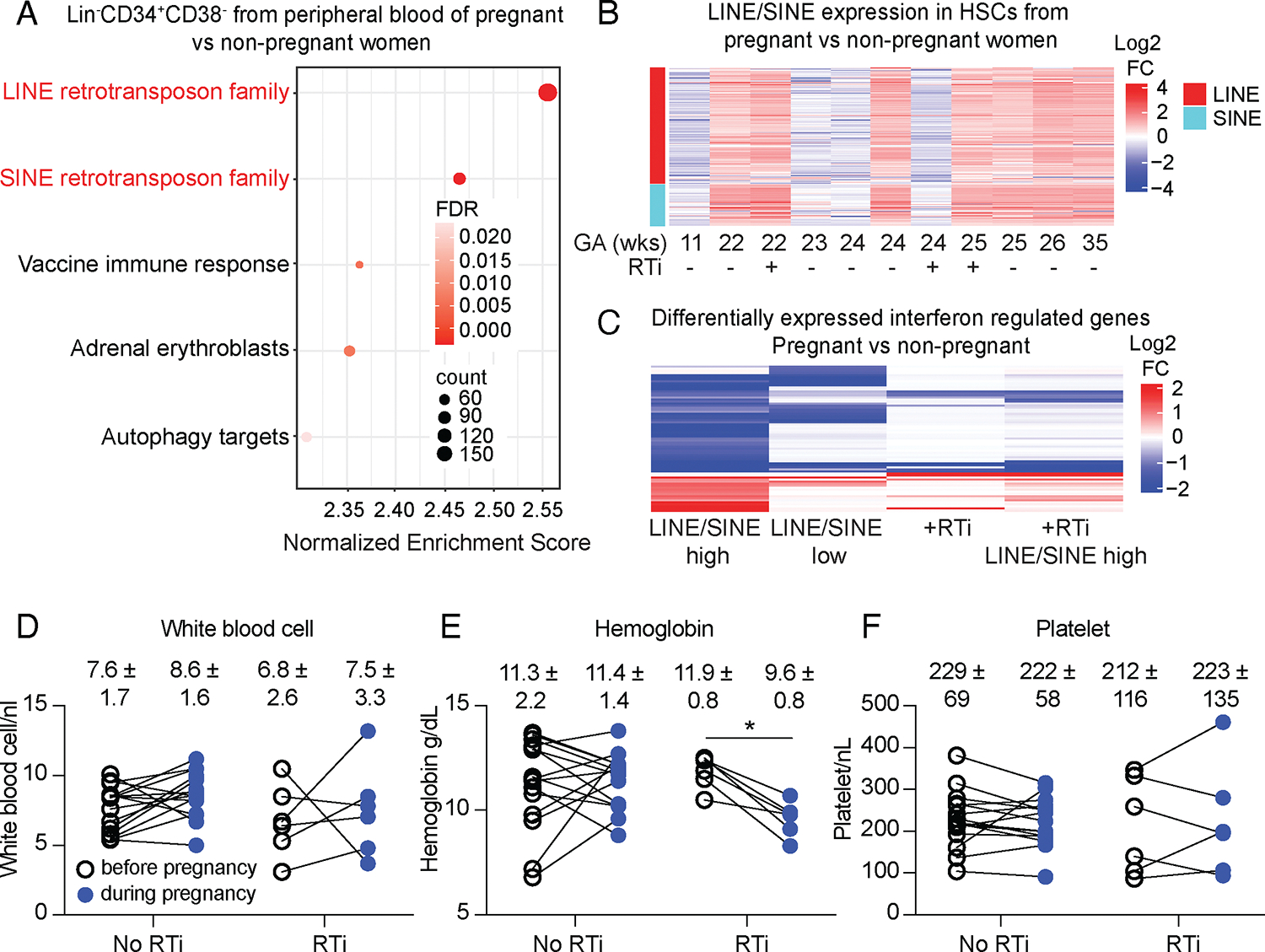
Hematopoietic stem cells (HSCs) and erythropoiesis are activated during pregnancy and after bleeding by the derepression of retrotransposons, including endogenous retroviruses and long interspersed nuclear elements. Retrotransposon transcription activates the innate immune sensors cyclic guanosine 3',5'-monophosphate-adenosine 5'-monophosphate synthase (cGAS) and stimulator of interferon (IFN) genes (STING), which induce IFN and IFN-regulated genes in HSCs, increasing HSC division and erythropoiesis. Inhibition of reverse transcriptase or deficiency for cGAS or STING had little or no effect on hematopoiesis in nonpregnant mice but depleted HSCs and erythroid progenitors in pregnant mice, reducing red blood cell counts. Retrotransposons and IFN-regulated genes were also induced in mouse HSCs after serial bleeding and, in human HSCs, during pregnancy. Reverse transcriptase inhibitor use was associated with anemia in pregnant but not in nonpregnant people, suggesting conservation of these mechanisms from mice to humans.
Conflict of interest statement
Figures
References
-
- Kickler TS, Spivak JL, Effect of repeated whole blood donations on serum immunoreactive erythropoietin levels in autologous donors. JAMA 260, 65–67 (1988). - PubMed
-
- Ebert BL, Bunn HF, Regulation of the erythropoietin gene. Blood 94, 1864–1877 (1999). - PubMed
-
- Cheshier SH, Prohaska SS, Weissman IL, The effect of bleeding on hematopoietic stem cell cycling and self-renewal. Stem Cells Dev 16, 707–717 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
