

Caspase-1 in Cx3cr1-expressing cells drives an IL-18-dependent T cell response that promotes parasite control during acute Toxoplasma gondii infection
- PMID: 39446964
- PMCID: PMC11537422
- DOI: 10.1371/journal.ppat.1012006
Caspase-1 in Cx3cr1-expressing cells drives an IL-18-dependent T cell response that promotes parasite control during acute Toxoplasma gondii infection
Abstract
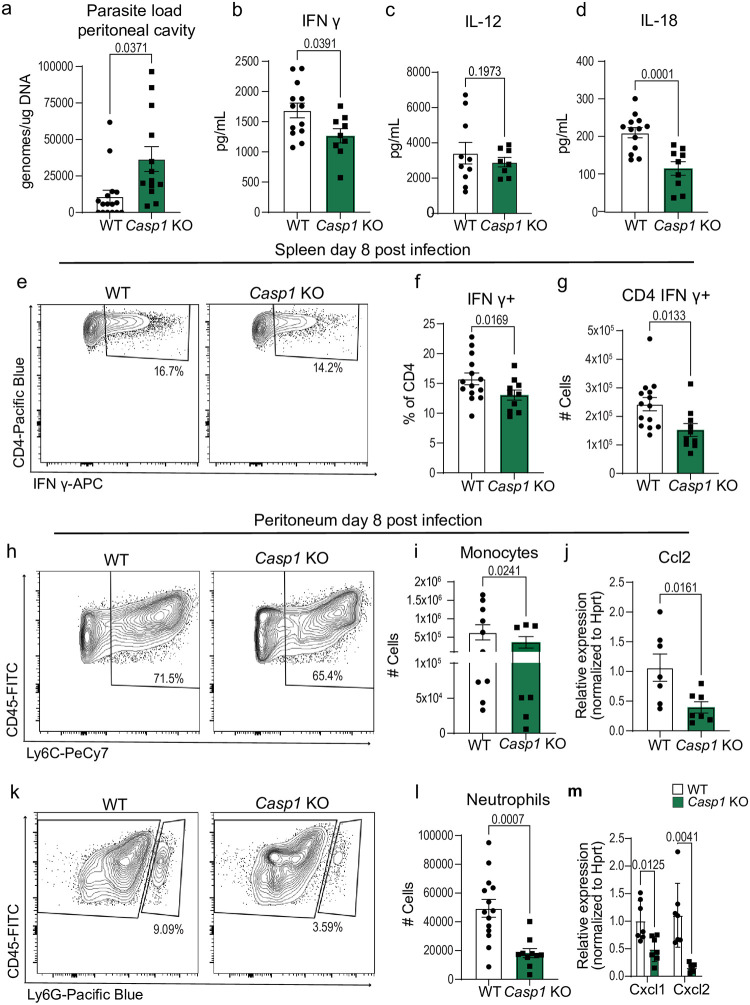
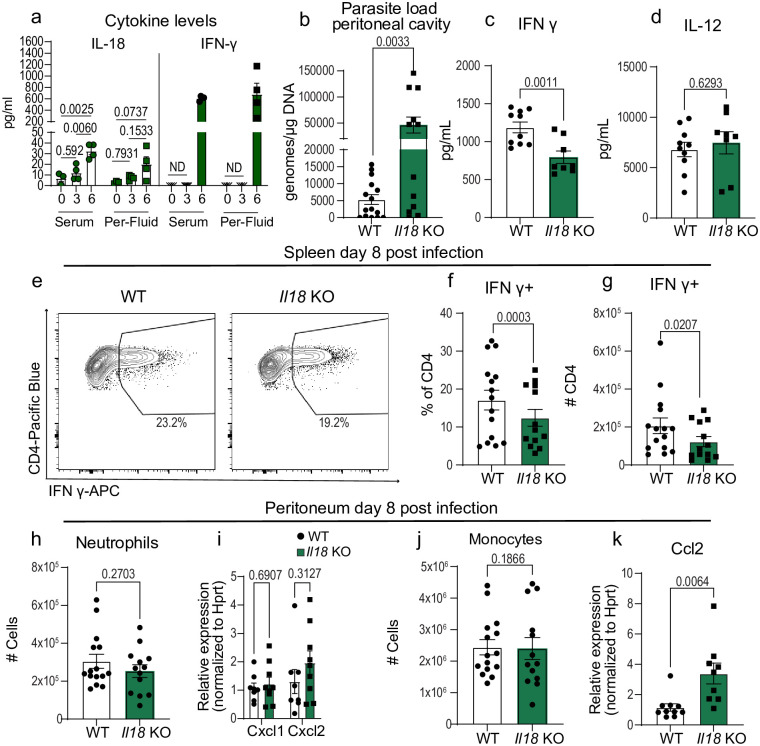
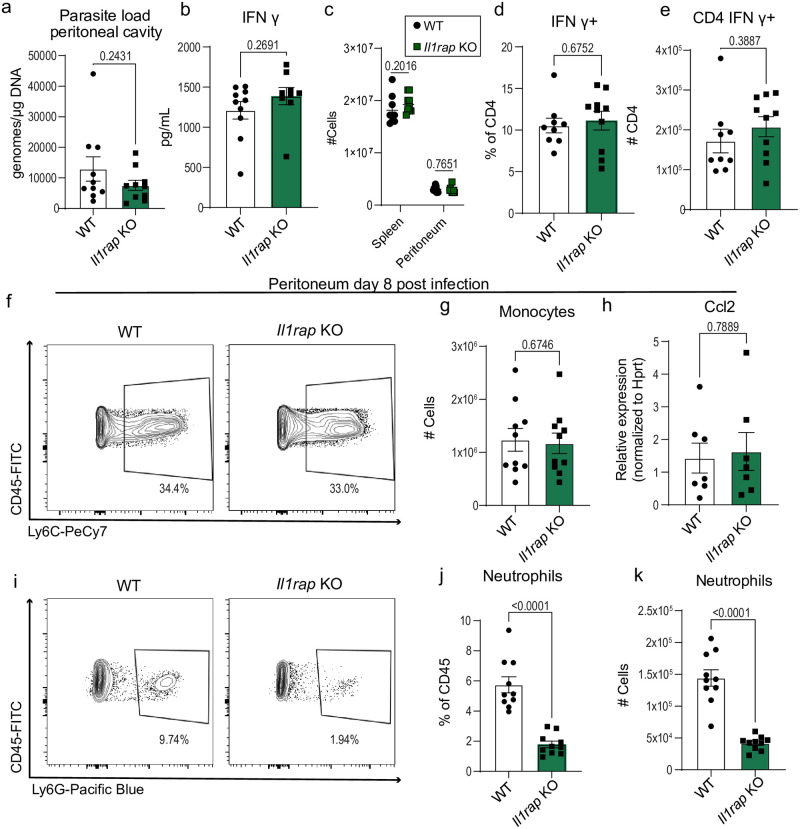
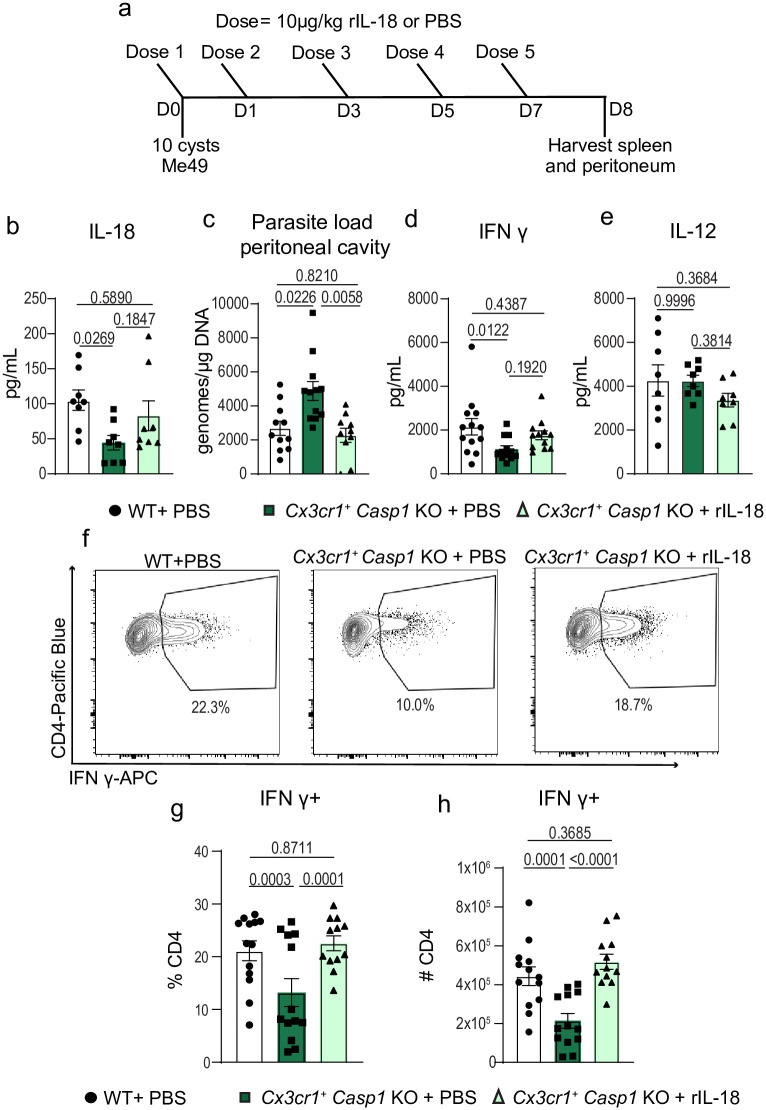
Inflammasome activation is a robust innate immune mechanism that promotes inflammatory responses through the release of alarmins and leaderless cytokines, including IL-1α, IL-1β, and IL-18. Various stimuli, including infectious agents and cellular stress, cause inflammasomes to assemble and activate caspase-1. Then, caspase-1 cleaves targets that lead to pore formation and leaderless cytokine activation and release. Toxoplasma gondii has been shown to promote inflammasome formation, but the cell types utilizing caspase-1 and the downstream effects on immunological outcomes during acute in vivo infection have not been explored. Here, using knockout mice, we examine the role of caspase-1 responses during acute T. gondii infection globally and in Cx3cr1-positive populations. We provide in vivo evidence that caspase-1 expression is critical for, IL-18 release, optimal interferon-γ (IFN-γ) production, monocyte and neutrophil recruitment to the site of infection, and parasite control. Specifically, we find that caspase-1 expression in Cx3cr1-positive cells drives IL-18 release, which potentiates CD4+ T cell IFN-γ production and parasite control. Notably, our Cx3cr1-Casp1 knockouts exhibited a selective T cell defect, mirroring the phenotype observed in Il18 knockouts. In further support of this finding, treatment of Cx3cr1-Casp1 knockout mice with recombinant IL-18 restored CD4+ T cell IFN-γ responses and parasite control. Additionally, we show that neutrophil recruitment is dependent on IL-1 receptor accessory protein (IL-1RAP) signaling but is dispensable for parasite control. Overall, these experiments highlight the multifaceted role of caspase-1 in multiple cell populations contributing to specific pathways that collectively contribute to caspase-1 dependent immunity to T. gondii.
Copyright: © 2024 Babcock et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Scharton-Kersten T, Contursi C, Masumi A, Sher A, Ozato K. Interferon consensus sequence binding protein-deficient mice display impaired resistance to intracellular infection due to a primary defect in interleukin 12 p40 induction. J Exp Med. 1997;186(9):1523–34. Epub 1997/11/14. doi: 10.1084/jem.186.9.1523 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
