

Alternative sulphur metabolism in the fungal pathogen Candida parapsilosis
- PMID: 39448588
- PMCID: PMC11502921
- DOI: 10.1038/s41467-024-53442-8
Alternative sulphur metabolism in the fungal pathogen Candida parapsilosis
Abstract
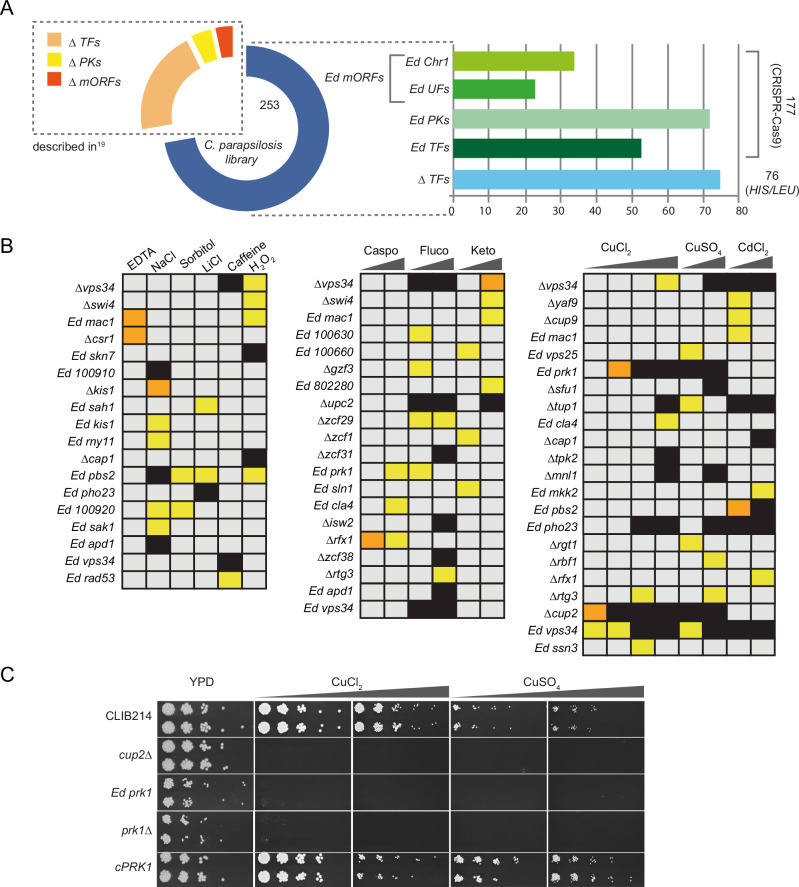
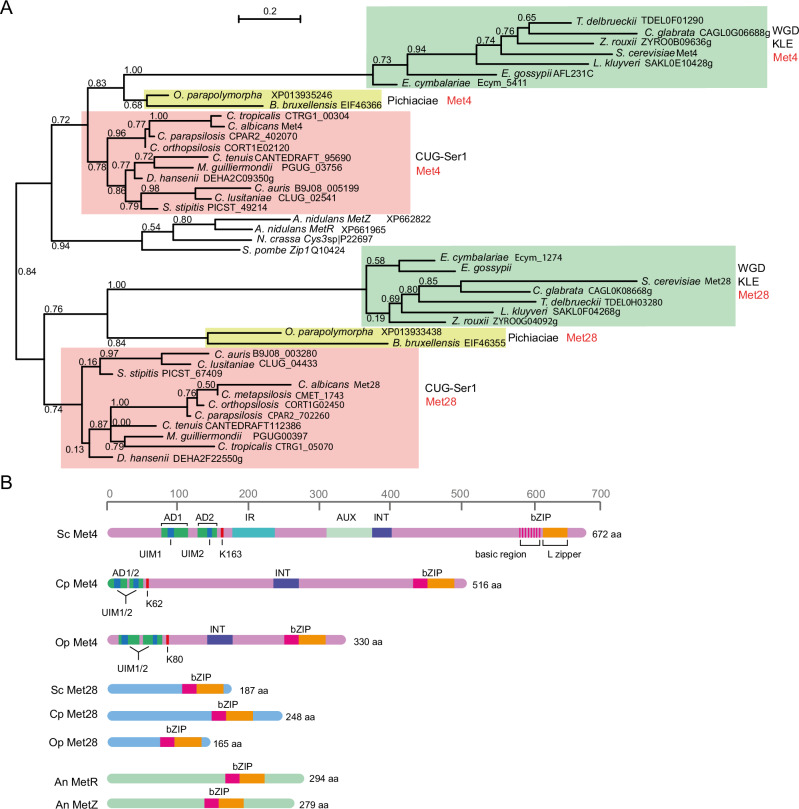
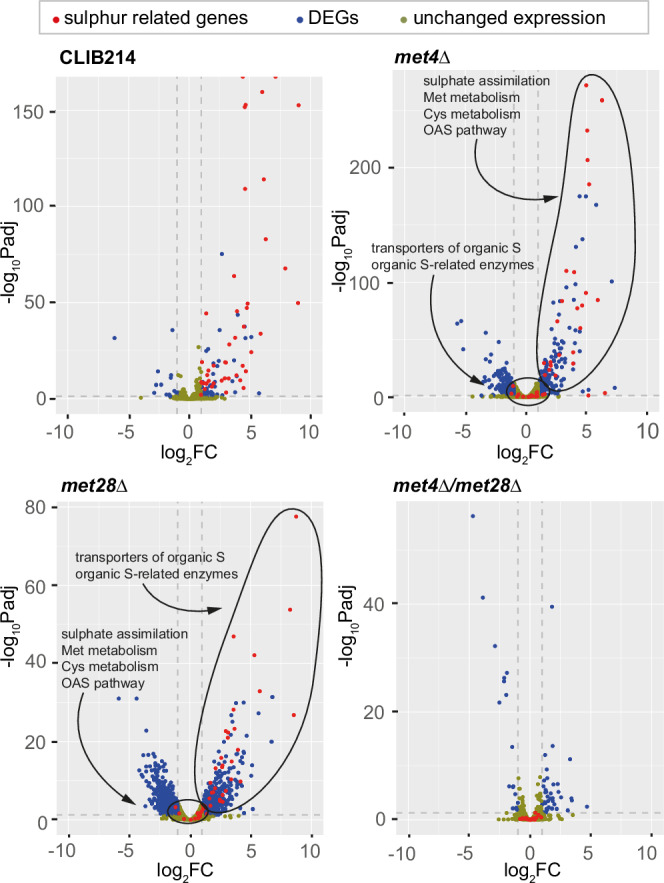
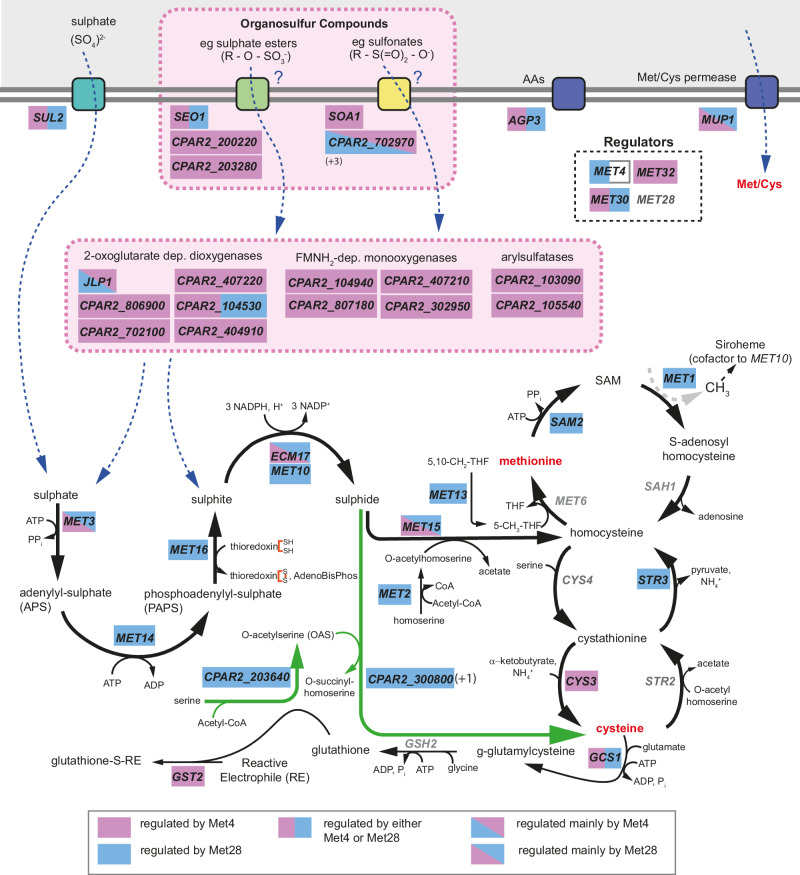
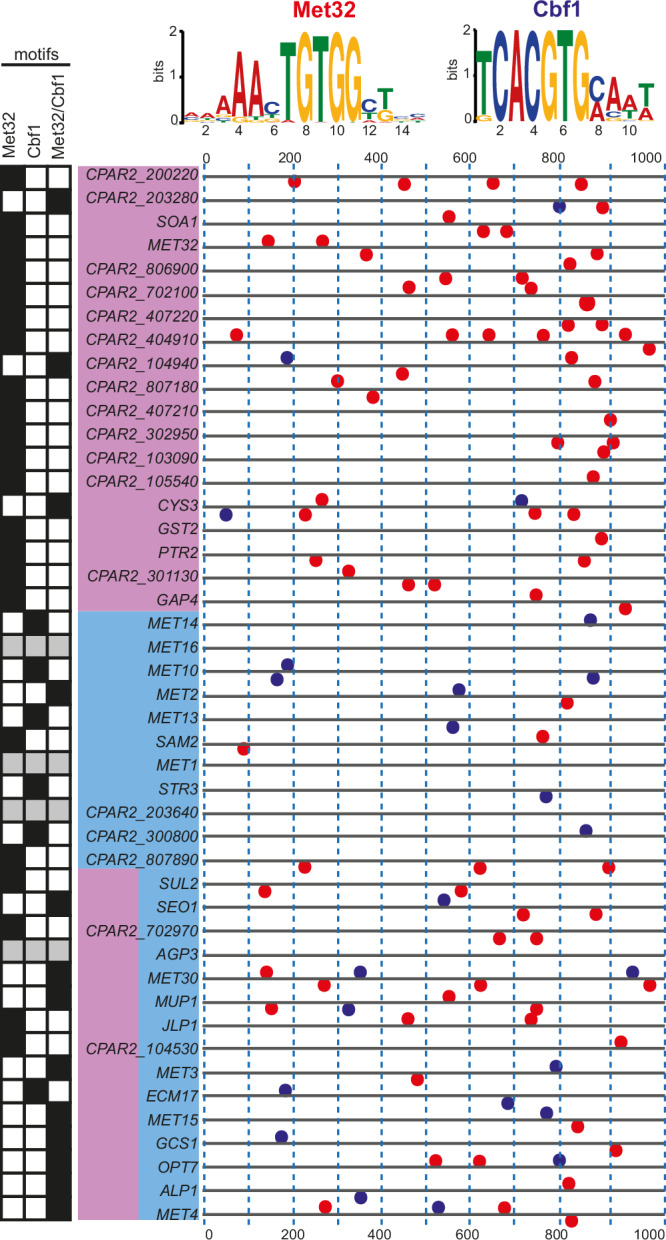
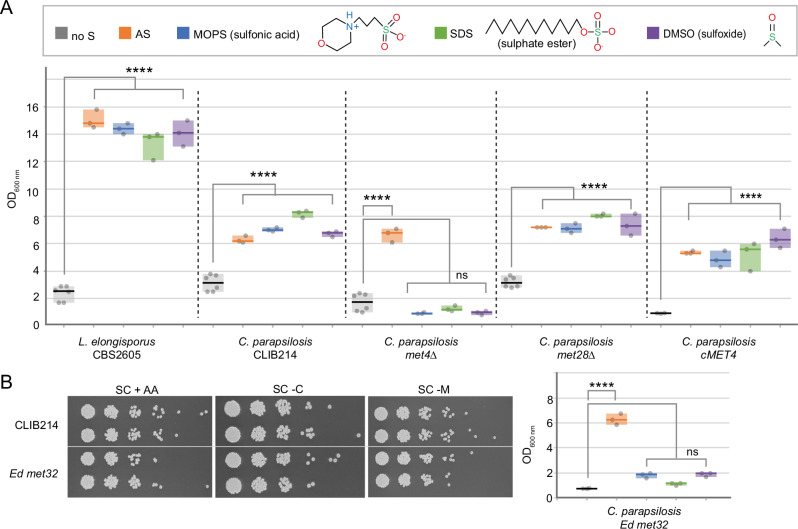
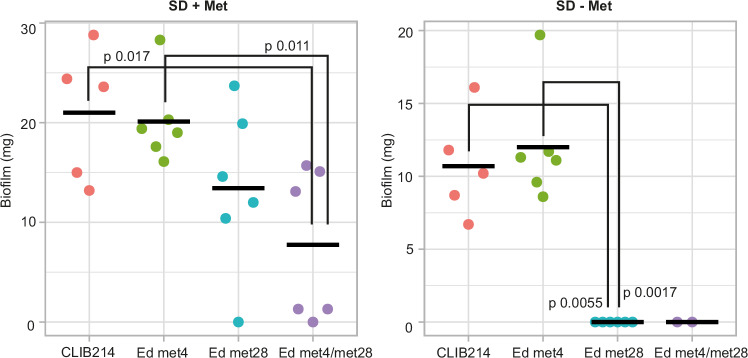
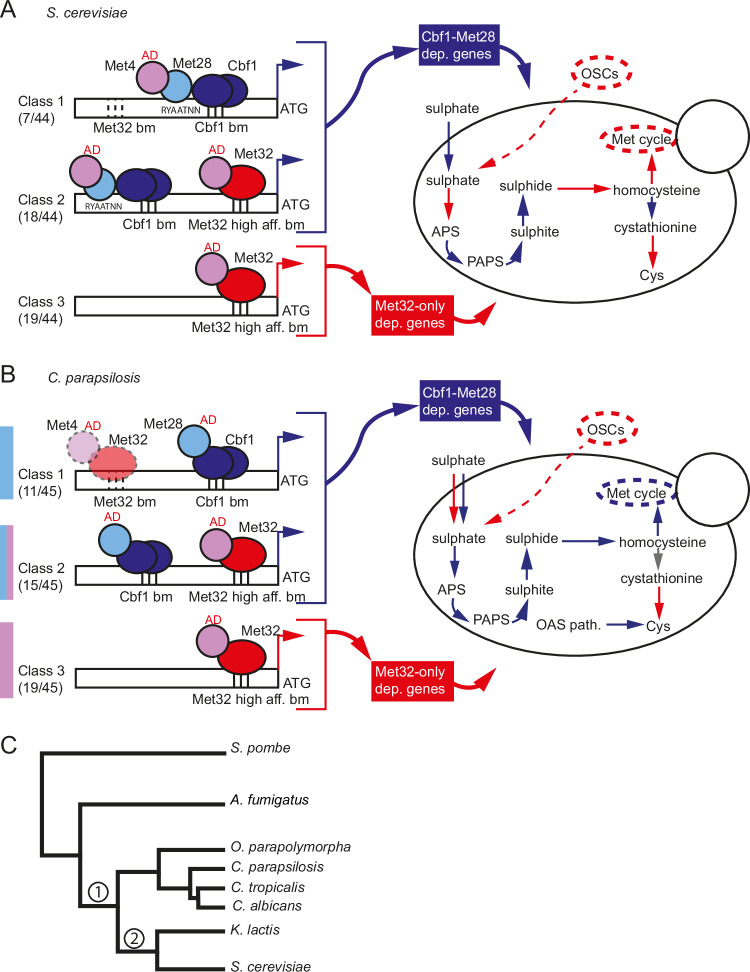
Candida parapsilosis is an opportunistic fungal pathogen commonly isolated from the environment and associated with nosocomial infection outbreaks worldwide. We describe here the construction of a large collection of gene disruptions, greatly increasing the molecular tools available for probing gene function in C. parapsilosis. We use these to identify transcription factors associated with multiple metabolic pathways, and in particular to dissect the network regulating the assimilation of sulphur. We find that, unlike in other yeasts and filamentous fungi, the transcription factor Met4 is not the main regulator of methionine synthesis. In C. parapsilosis, assimilation of inorganic sulphur (sulphate) and synthesis of cysteine and methionine is regulated by Met28, a paralog of Met4, whereas Met4 regulates expression of a wide array of transporters and enzymes involved in the assimilation of organosulfur compounds. Analysis of transcription factor binding sites suggests that Met4 is recruited by the DNA-binding protein Met32, and Met28 is recruited by Cbf1. Despite having different target genes, Met4 and Met28 have partial functional overlap, possibly because Met4 can contribute to assimilation of inorganic sulphur in the absence of Met28.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
