

Constitutive activation of two-component systems reveals regulatory network interactions in Streptococcus agalactiae
- PMID: 39448655
- PMCID: PMC11502775
- DOI: 10.1038/s41467-024-53439-3
Constitutive activation of two-component systems reveals regulatory network interactions in Streptococcus agalactiae
Abstract
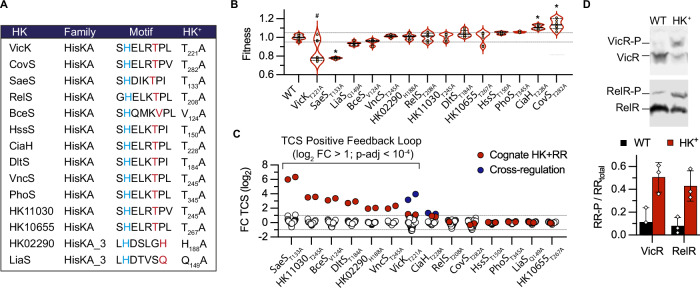
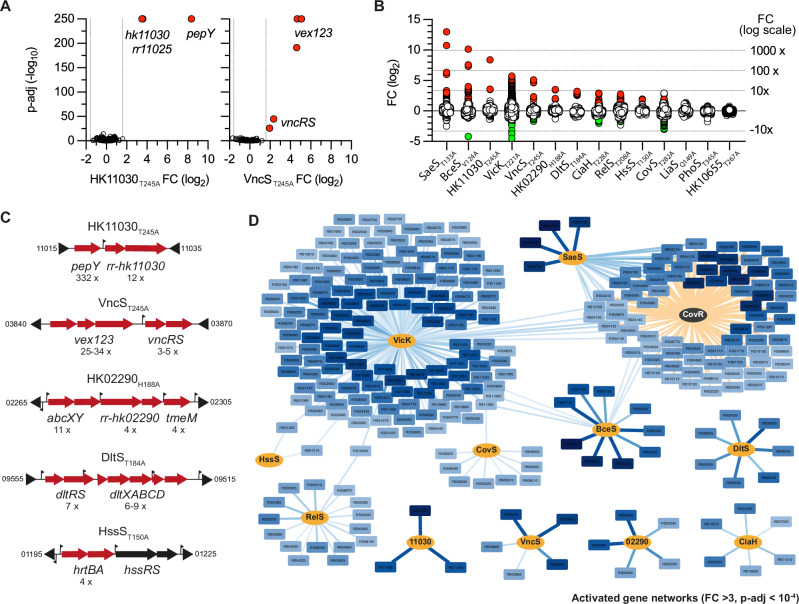
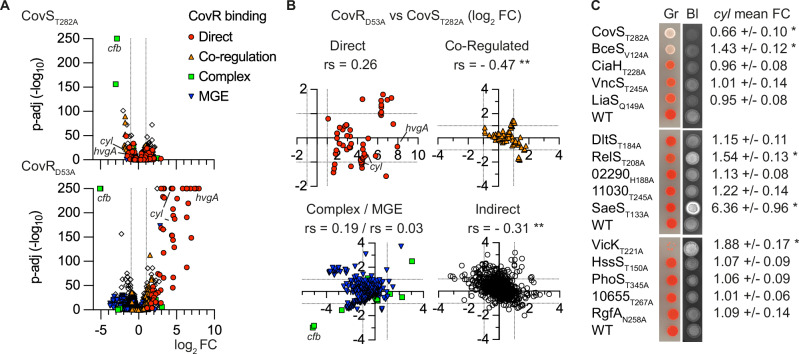
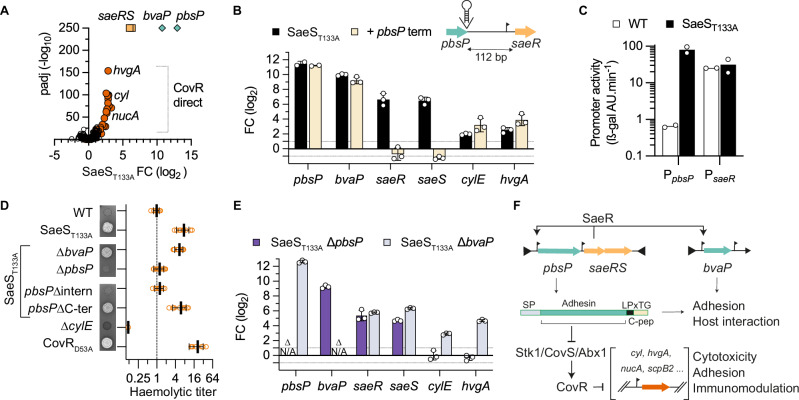
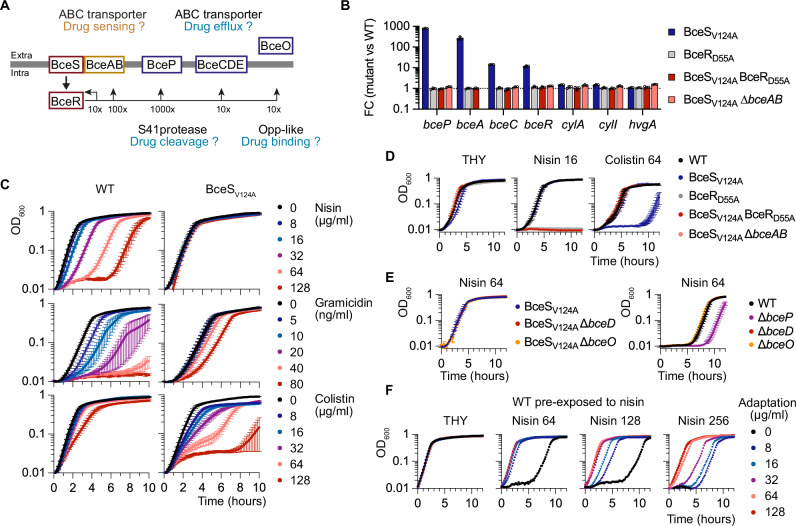
Bacterial two-component systems (TCSs) are signaling modules that control physiology, adaptation, and host interactions. A typical TCS consists of a histidine kinase (HK) that activates a response regulator via phosphorylation in response to environmental signals. Here, we systematically test the effect of inactivating the conserved phosphatase activity of HKs to activate TCS signaling pathways. Transcriptome analyses of 14 HK mutants in Streptococcus agalactiae, the leading cause of neonatal meningitis, validate the conserved HK phosphatase mechanism and its role in the inhibition of TCS activity in vivo. Constitutive TCS activation, independent of environmental signals, enables high-resolution mapping of the regulons for several TCSs (e.g., SaeRS, BceRS, VncRS, DltRS, HK11030, HK02290) and reveals the functional diversity of TCS signaling pathways, ranging from highly specialized to interconnected global regulatory networks. Targeted analysis shows that the SaeRS-regulated PbsP adhesin acts as a signaling molecule to activate CovRS signaling, thereby linking the major regulators of host-pathogen interactions. Furthermore, constitutive BceRS activation reveals drug-independent activity, suggesting a role in cell envelope homeostasis beyond antimicrobial resistance. This study highlights the versatility of constitutive TCS activation, via phosphatase-deficient HKs, to uncover regulatory networks and biological processes.
© 2024. The Author(s).
Conflict of interest statement
GT is an employee and CB is the founder and owner of Scylla Biotech Srl. The company did not provide funding and had no role in the design, conduct, or publication of the study. All other authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
