

In Vivo Biocompatibility of Synechococcus sp. PCC 7002-Integrated Scaffolds for Skin Regeneration
- PMID: 39452593
- PMCID: PMC11508603
- DOI: 10.3390/jfb15100295
In Vivo Biocompatibility of Synechococcus sp. PCC 7002-Integrated Scaffolds for Skin Regeneration
Abstract
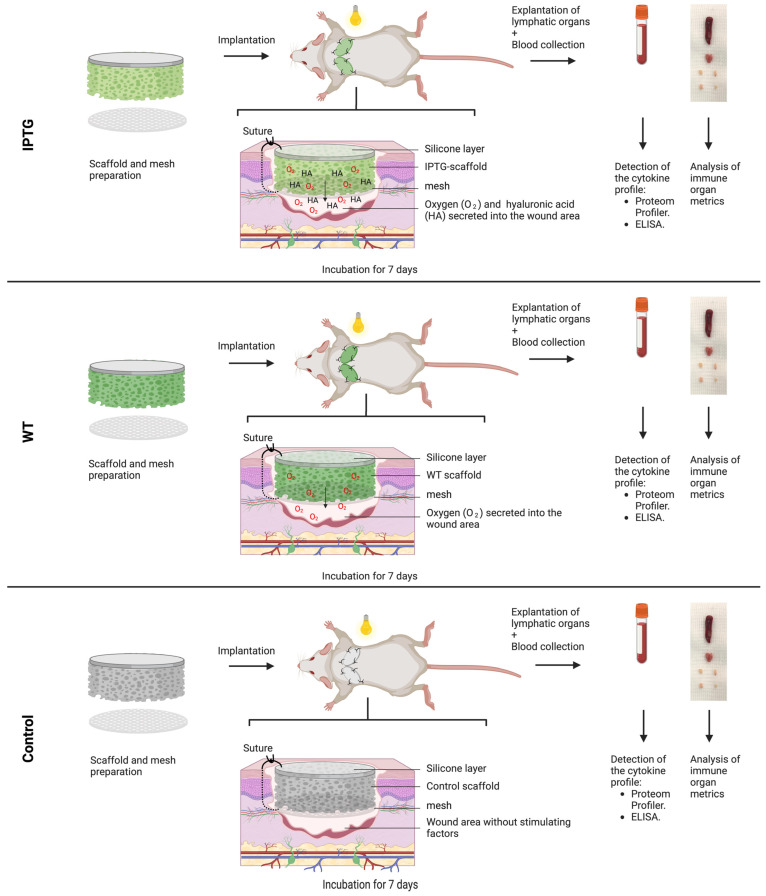
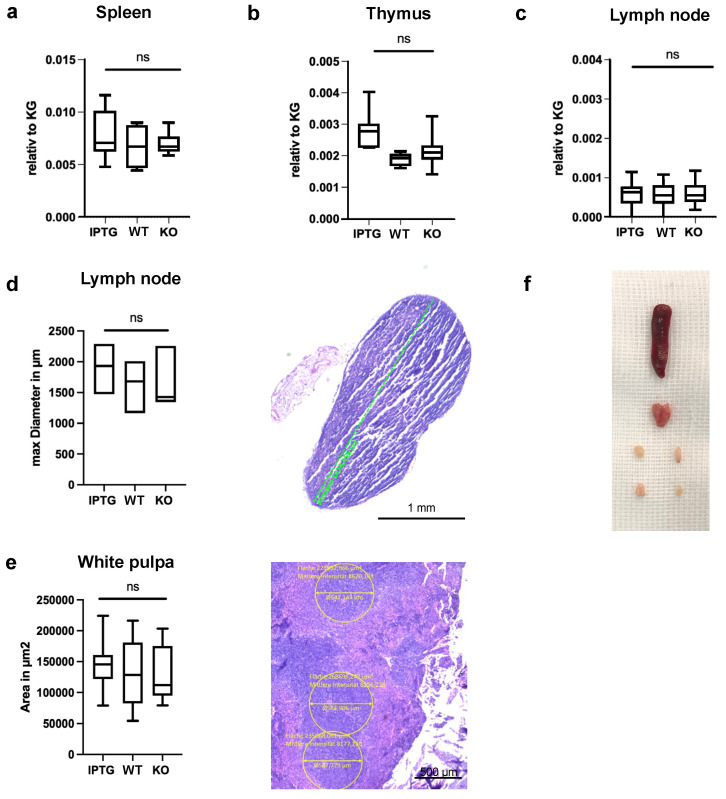
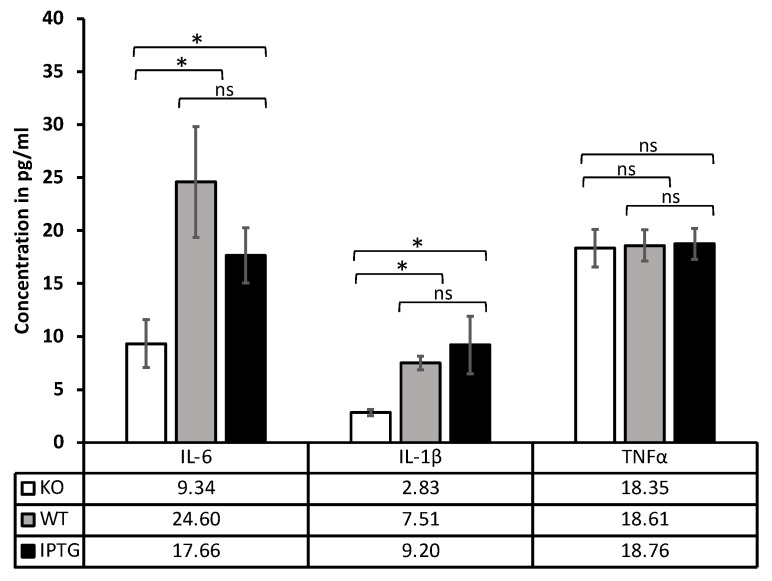
Cyanobacteria, commonly known as blue-green algae, are prevalent in freshwater systems and have gained interest for their potential in medical applications, particularly in skin regeneration. Among these, Synechococcus sp. strain PCC 7002 stands out because of its rapid proliferation and capacity to be genetically modified to produce growth factors. This study investigates the safety of Synechococcus sp. PCC 7002 when used in scaffolds for skin regeneration, focusing on systemic inflammatory responses in a murine model. We evaluated the following three groups: scaffolds colonized with genetically engineered bacteria producing hyaluronic acid, scaffolds with wild-type bacteria, and control scaffolds without bacteria. After seven days, we assessed systemic inflammation by measuring changes in cytokine profiles and lymphatic organ sizes. The results showed no significant differences in spleen, thymus, and lymph node weights, indicating a lack of overt systemic toxicity. Blood cytokine analysis revealed elevated levels of IL-6 and IL-1β in scaffolds with bacteria, suggesting a systemic inflammatory response, while TNF-α levels remained unaffected. Proteome profiling identified distinct cytokine patterns associated with bacterial colonization, including elevated inflammatory proteins and products, indicative of acute inflammation. Conversely, control scaffolds exhibited protein profiles suggestive of a rejection response, characterized by increased levels of cytokines involved in T and B cell activation. Our findings suggest that Synechococcus sp. PCC 7002 does not appear to cause significant systemic toxicity, supporting its potential use in biomedical applications. Further research is necessary to explore the long-term effects and clinical implications of these responses.
Keywords: Synechococcus sp. PCC 7002; biocompatibility; biomaterials; cyanobacteria; scaffolds; tissue engineering.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures

References
-
- Sarnaik A., Abernathy M.H., Han X., Ouyang Y., Xia K., Chen Y., Cress B., Zhang F., Lali A., Pandit R., et al. Metabolic engineering of cyanobacteria for photoautotrophic production of heparosan, a pharmaceutical precursor of heparin. Algal Res. 2019;37:57–63. doi: 10.1016/j.algal.2018.11.010. - DOI
LinkOut - more resources
Full Text Sources
