

Biallelic germline DDX41 variants in a patient with bone dysplasia, ichthyosis, and dysmorphic features
- PMID: 39453476
- PMCID: PMC11576897
- DOI: 10.1007/s00439-024-02708-8
Biallelic germline DDX41 variants in a patient with bone dysplasia, ichthyosis, and dysmorphic features
Abstract
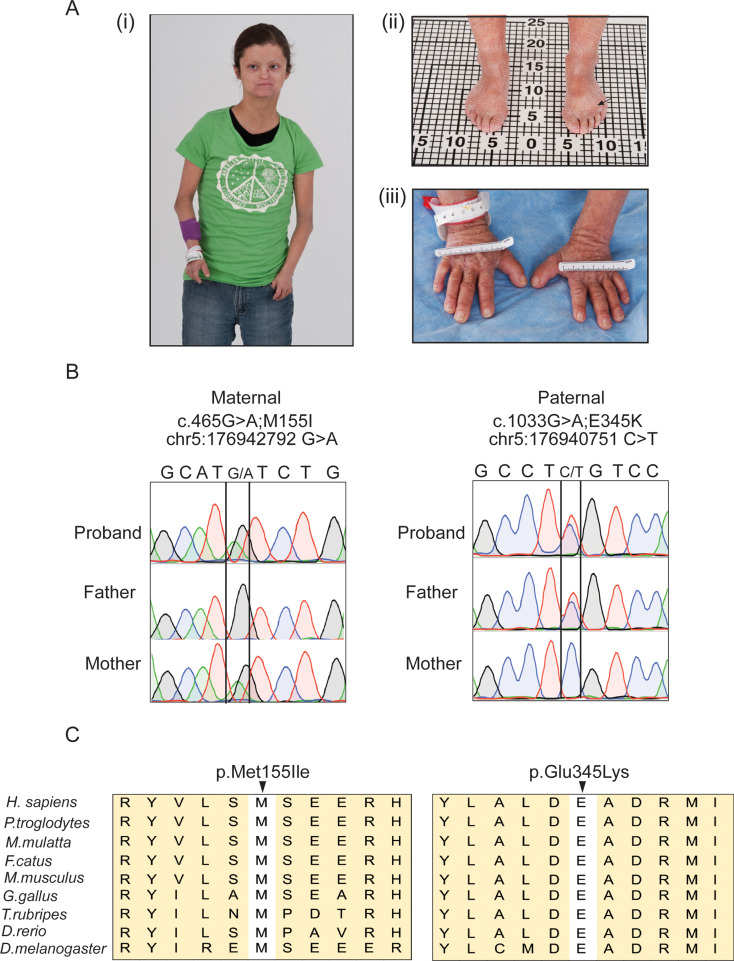
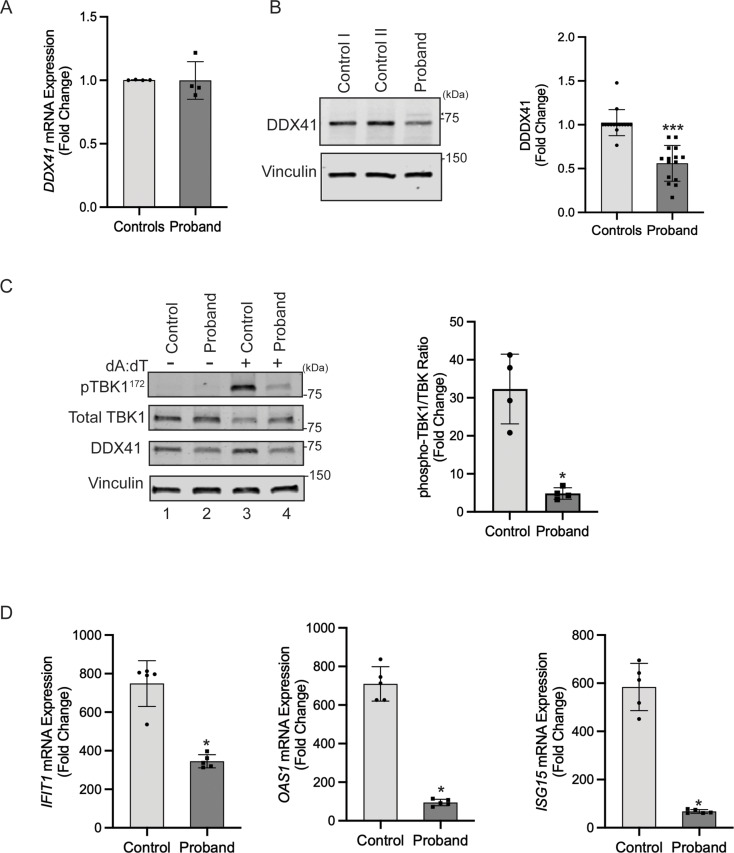
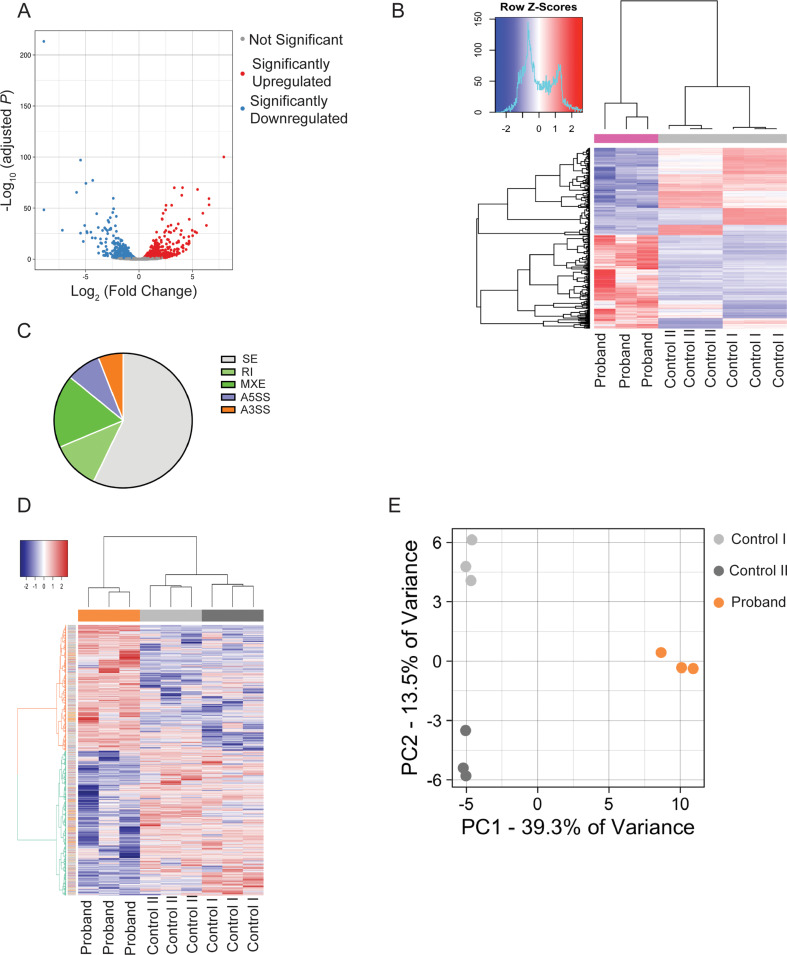
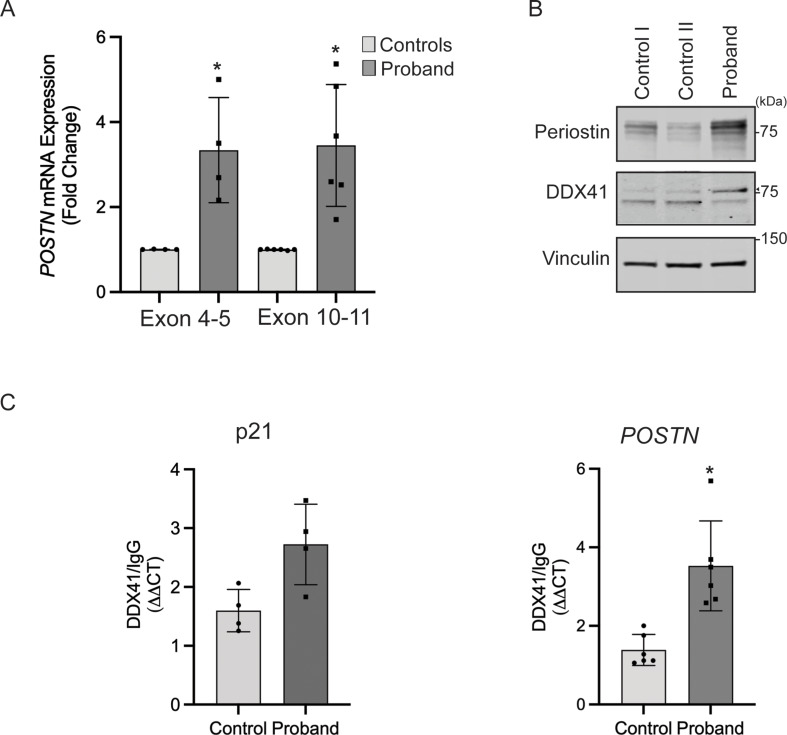
DDX41 (DEAD‑box helicase 41) is a member of the largest family of RNA helicases. The DEAD-box RNA helicases share a highly conserved core structure and regulate all aspects of RNA metabolism. The functional role of DDX41 in innate immunity is also highly conserved. DDX41 acts as a sensor of viral DNA and activates the STING-TBK1-IRF3-type I IFN signaling pathway. Germline heterozygous variants in DDX41 have been reported in familial myelodysplasia syndrome (MDS)/acute myeloid leukemia (AML) patients; most patients also acquired a somatic variant in the second DDX41 allele. Here, we report a patient who inherited compound heterozygous DDX41 variants and presented with bone dysplasia, ichthyosis, and dysmorphic features. Functional analyses of the patient-derived dermal fibroblasts revealed a reduced abundance of DDX41 and abrogated activation of the IFN genes through the STING-type I interferon pathway. Genome-wide transcriptome analyses in the patient's fibroblasts revealed significant gene dysregulation and changes in the RNA splicing events. The patient's fibroblasts also displayed upregulation of periostin mRNA expression. Using an RNA binding protein assay, we identified DDX41 as a novel regulator of periostin expression. Our results suggest that functional impairment of DDX41, along with dysregulated periostin expression, likely contributes to this patient's multisystem disorder.
© 2024. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
Figures
References
-
- Chlon TM, Stepanchick E, Hershberger CE, Daniels NJ, Hueneman KM, Kuenzi Davis A, Choi K, Zheng Y, Gurnari C, Haferlach T, Padgett RA, Maciejewski JP, Starczynowski DT (2021) Germline DDX41 mutations cause ineffective hematopoiesis and myelodysplasia. Cell Stem Cell 28:1966–1981 e6. 10.1016/j.stem.2021.08.004 - DOI - PMC - PubMed
-
- Gahl WA, Markello TC, Toro C, Fajardo KF, Sincan M, Gill F, Carlson-Donohoe H, Gropman A, Pierson TM, Golas G, Wolfe L, Groden C, Godfrey R, Nehrebecky M, Wahl C, Landis DM, Yang S, Madeo A, Mullikin JC, Boerkoel CF, Tifft CJ, Adams D (2012) The National Institutes of Health Undiagnosed diseases Program: insights into rare diseases. Genet Med 14:51–59. 10.1038/gim.0b013e318232a005 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
