

Isolated theta waves originating from the midline thalamus trigger memory reactivation during NREM sleep in mice
- PMID: 39455583
- PMCID: PMC11511994
- DOI: 10.1038/s41467-024-53522-9
Isolated theta waves originating from the midline thalamus trigger memory reactivation during NREM sleep in mice
Abstract
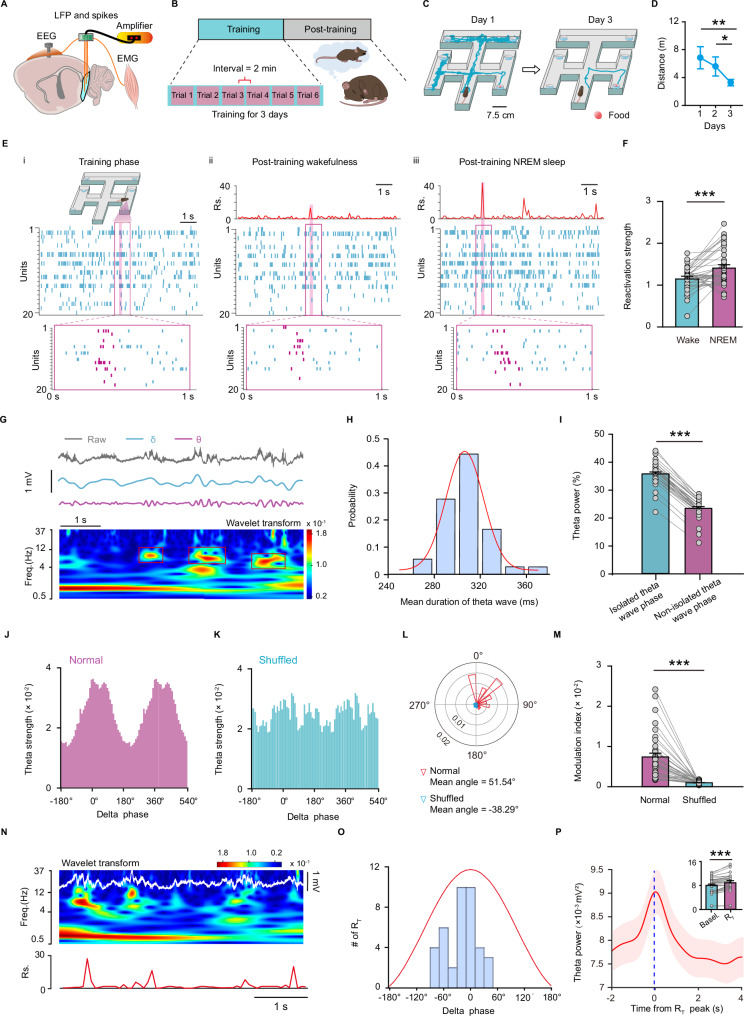
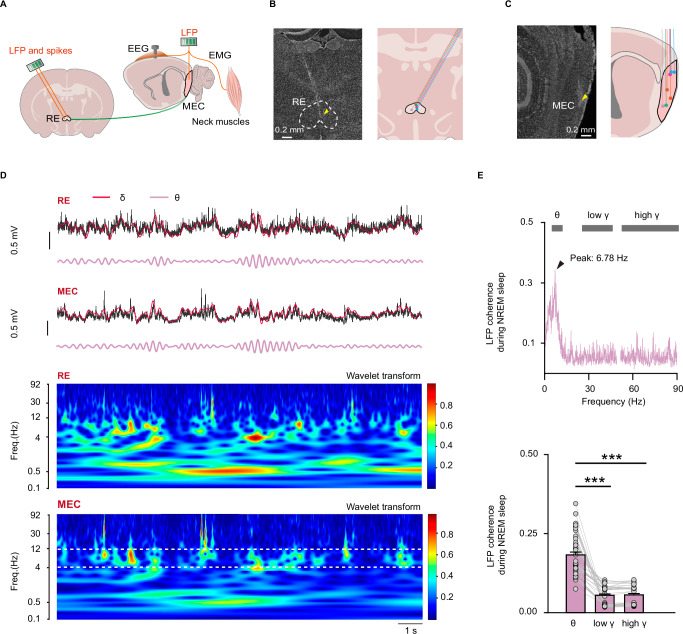
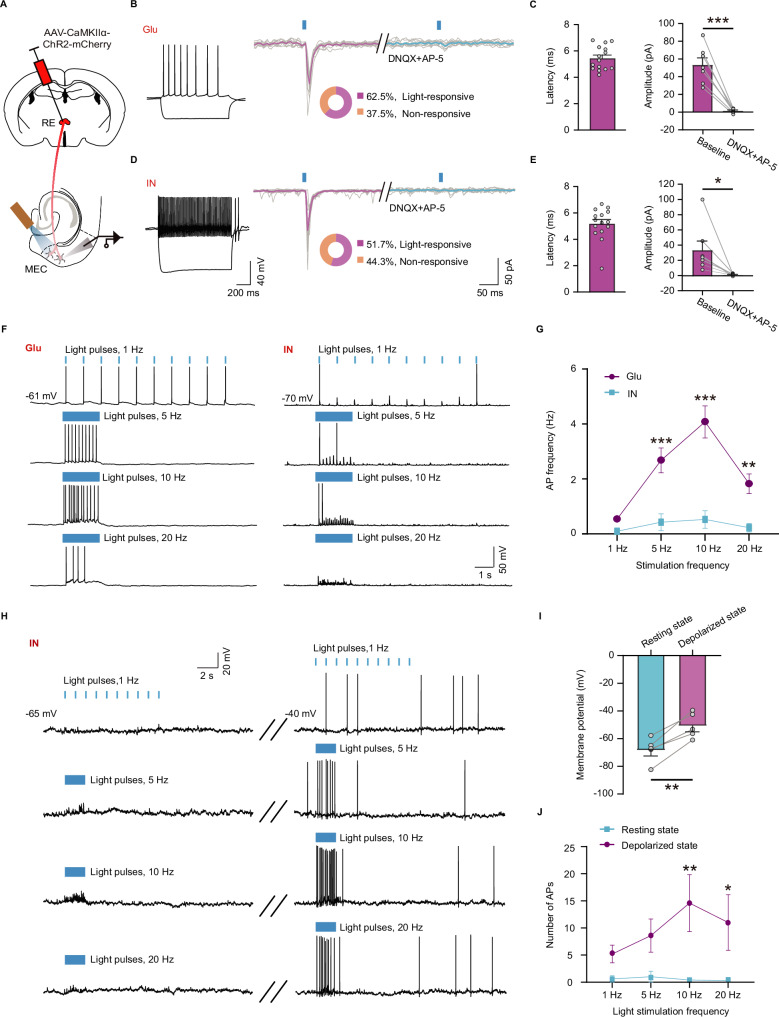
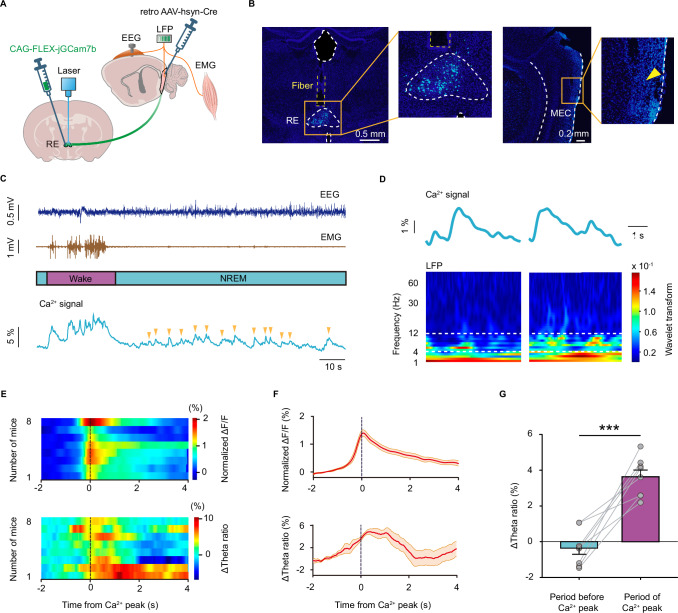
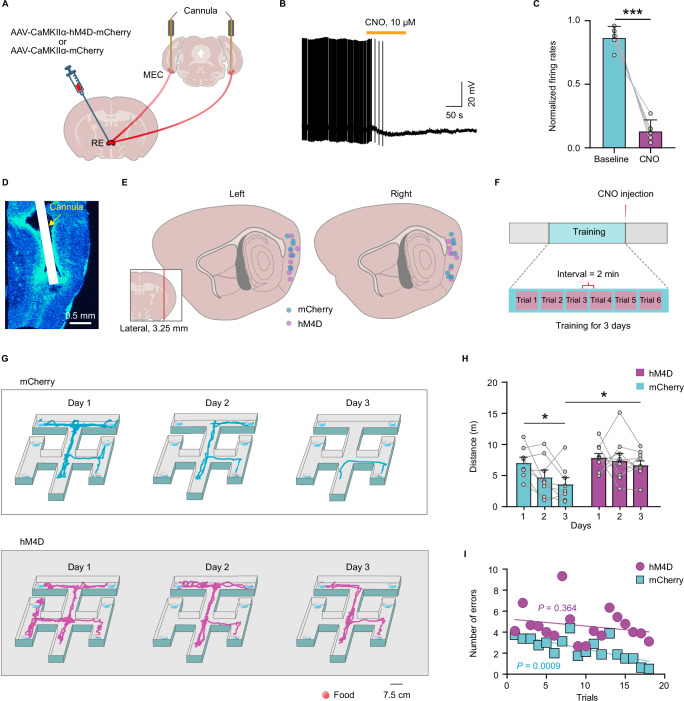
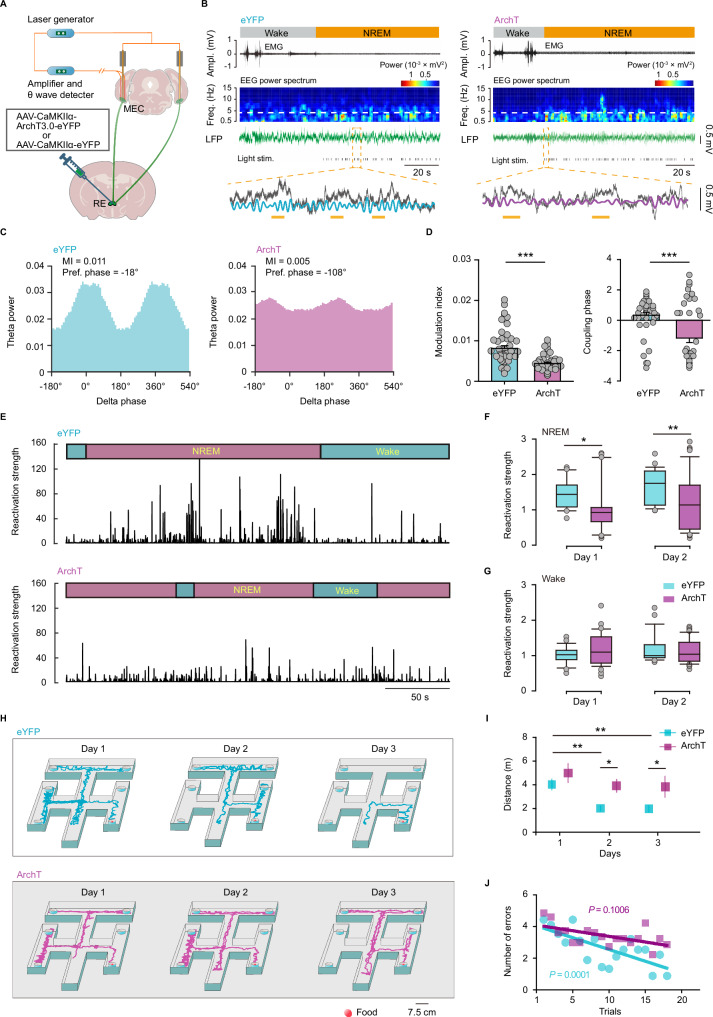
During non-rapid eye movement (NREM) sleep, neural ensembles in the entorhinal-hippocampal circuit responsible for encoding recent memories undergo reactivation to facilitate the process of memory consolidation. This reactivation is widely acknowledged as pivotal for the formation of stable memory and its impairment is closely associated with memory dysfunction. To date, the neural mechanisms driving the reactivation of neural ensembles during NREM sleep remain poorly understood. Here, we show that the neural ensembles in the medial entorhinal cortex (MEC) that encode spatial experiences exhibit reactivation during NREM sleep. Notably, this reactivation consistently coincides with isolated theta waves. In addition, we found that the nucleus reuniens (RE) in the midline thalamus exhibits typical theta waves during NREM sleep, which are highly synchronized with those occurring in the MEC in male mice. Closed-loop optogenetic inhibition of the RE-MEC pathway specifically suppressed these isolated theta waves, resulting in impaired reactivation and compromised memory consolidation following a spatial memory task in male mice. The findings suggest that theta waves originating from the ventral midline thalamus play a role in initiating memory reactivation and consolidation during sleep.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
- 2021ZD0203201/Ministry of Science and Technology of the People's Republic of China (Chinese Ministry of Science and Technology)
- 2021ZD0203400/Ministry of Science and Technology of the People's Republic of China (Chinese Ministry of Science and Technology)
- 32371038/National Natural Science Foundation of China (National Science Foundation of China)
- 31921003/National Natural Science Foundation of China (National Science Foundation of China)
LinkOut - more resources
Full Text Sources
