

NRF2 and Thioredoxin Reductase 1 as Modulators of Interactions between Zinc and Selenium
- PMID: 39456464
- PMCID: PMC11505002
- DOI: 10.3390/antiox13101211
NRF2 and Thioredoxin Reductase 1 as Modulators of Interactions between Zinc and Selenium
Abstract
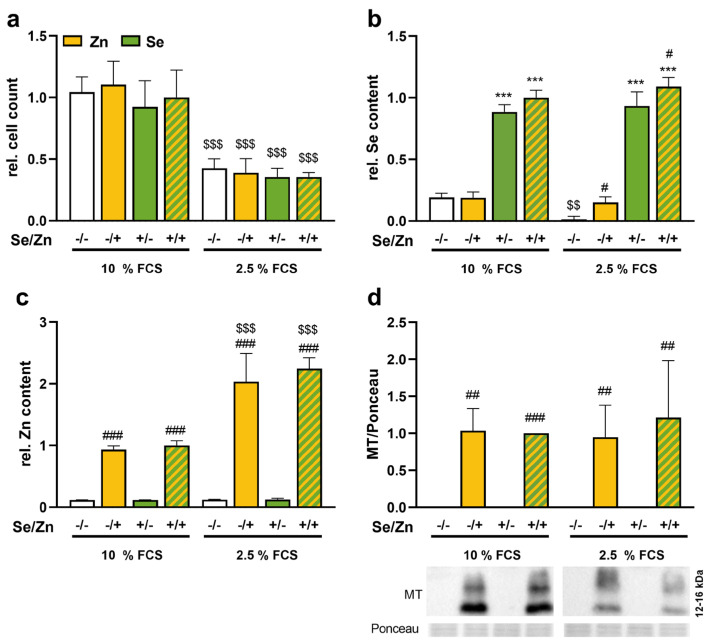
Background: Selenium and zinc are essential trace elements known to regulate cellular processes including redox homeostasis. During inflammation, circulating selenium and zinc concentrations are reduced in parallel, but underlying mechanisms are unknown. Accordingly, we modulated the zinc and selenium supply of HepG2 cells to study their relationship.
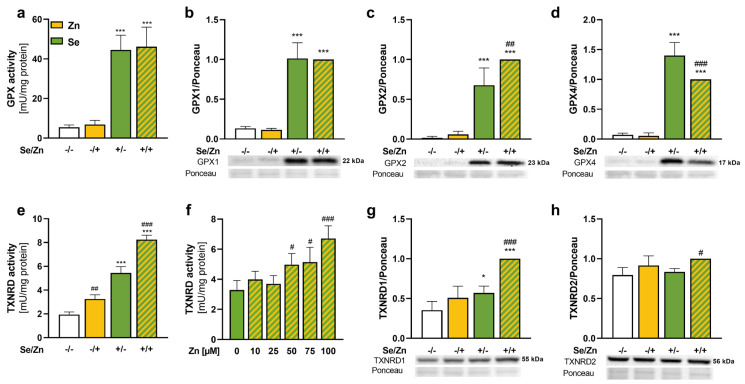
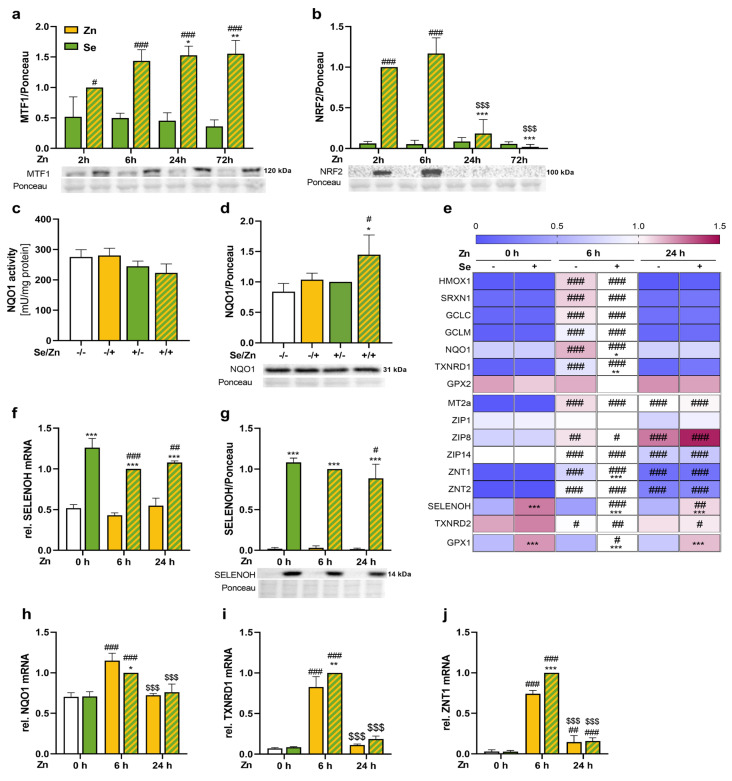
Methods: HepG2 cells were supplied with selenite in combination with a short- or long-term zinc treatment to investigate intracellular concentrations of selenium and zinc together with biomarkers describing their status. In addition, the activation of the redox-sensitive transcription factor NRF2 was analyzed.
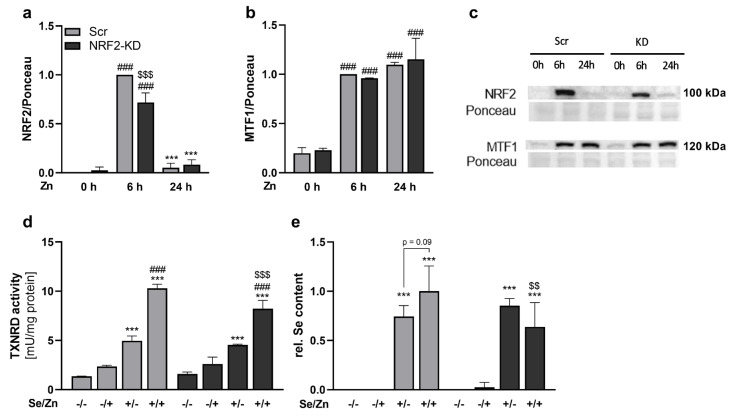
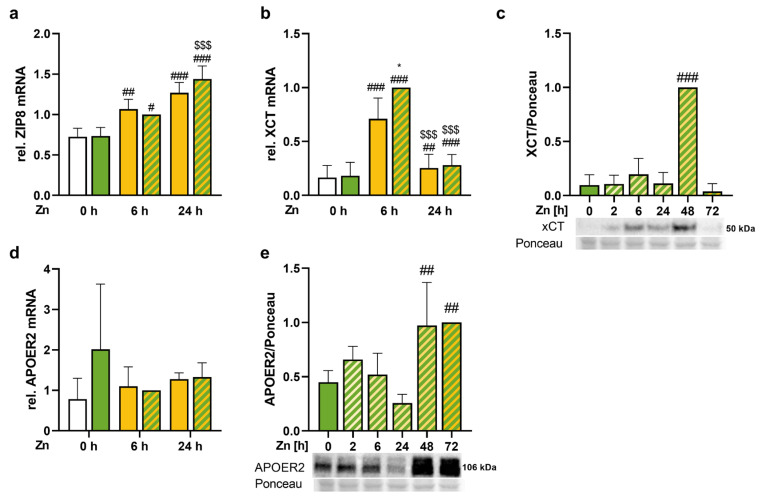
Results: Zinc not only increased the nuclear translocation of NRF2 after 2 to 6 h but also enhanced the intracellular selenium content after 72 h, when the cells were exposed to both trace elements. In parallel, the activity and expression of the selenoprotein thioredoxin reductase 1 (TXNRD1) increased, while the gene expression of other selenoproteins remained unaffected or was even downregulated. The zinc effects on the selenium concentration and TXNRD activity were reduced in cells with stable NRF2 knockdown in comparison to control cells.
Conclusions: This indicates a functional role of NRF2 in mediating the zinc/selenium crosstalk and provides an explanation for the observed unidirectional behavior of selenium and zinc.
Keywords: NRF2; selenium; selenium transporter; thioredoxin reductase; zinc.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Peters K.M., Galinn S.E., Tsuji P.A. Selenium: Dietary Sources, Human Nutritional Requirements and Intake across Populations. In: Hatfield D., Schweizer U., Tsuji P., Gladyshev V., editors. Selenium: Its Molecular Biology and Role in Human Health. Springer International Publishing; Cham, Switzerland: 2016. pp. 295–305.
Grants and funding
LinkOut - more resources
Full Text Sources
