

Modulation of ADAM17 Levels by Pestiviruses Is Species-Specific
- PMID: 39459898
- PMCID: PMC11512297
- DOI: 10.3390/v16101564
Modulation of ADAM17 Levels by Pestiviruses Is Species-Specific
Abstract
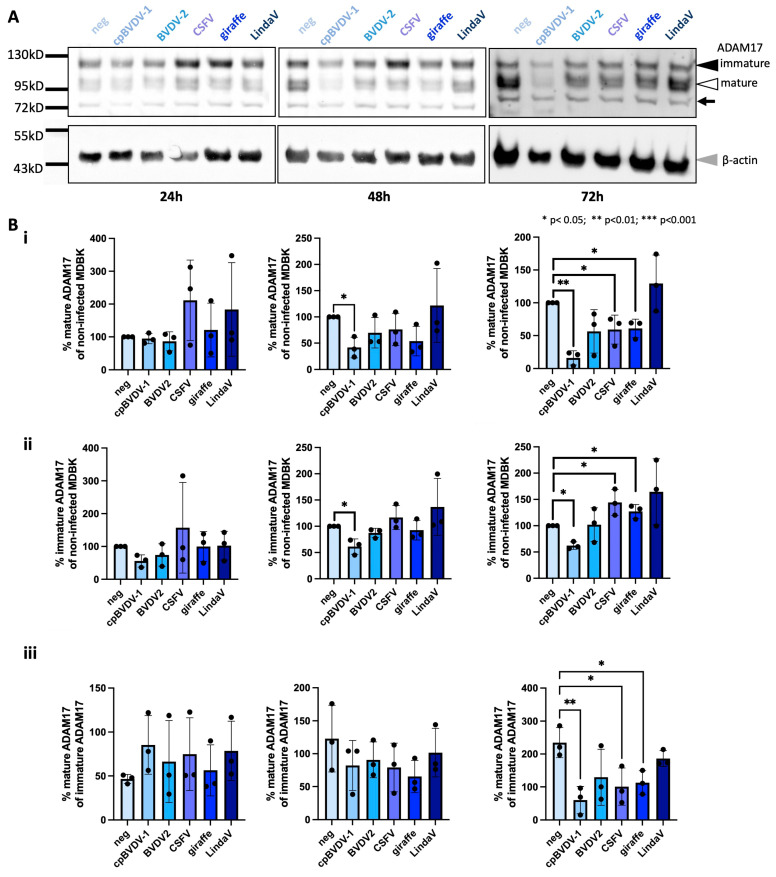
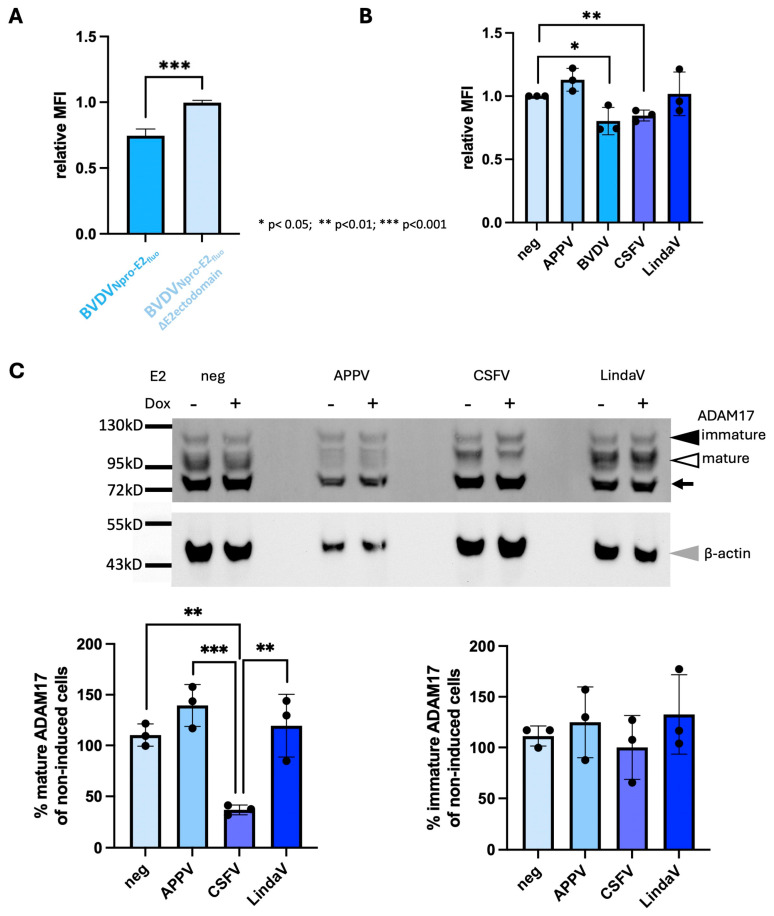
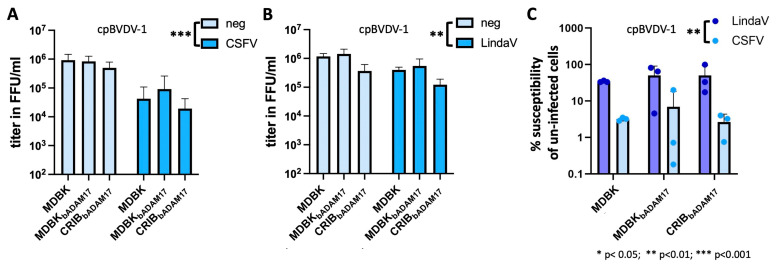
Upon host cell infection, viruses modulate their host cells to better suit their needs, including the downregulation of virus entry receptors. ADAM17, a cell surface sheddase, is an essential factor for infection of bovine cells with several pestiviruses. To assess the effect of pestivirus infection on ADAM17, the amounts of cellular ADAM17 and its presence at the cell surface were determined. Mature ADAM17 levels were reduced upon infection with a cytopathic pestivirus bovis (bovine viral diarrhea virus, cpBVDV), pestivirus suis (classical swine fever virus, CSFV) or pestivirus giraffae (strain giraffe), but not negatively affected by pestivirus L (Linda virus, LindaV). A comparable reduction of ADAM17 surface levels, which represents the bioactive form, could be observed in the presence of E2 of BVDV and CSFV, but not LindaV or atypical porcine pestivirus (pestivirus scrofae) E2. Superinfection exclusion in BVDV infection is caused by at least two proteins, glycoprotein E2 and protease/helicase NS3. To evaluate whether the lowered ADAM17 levels could be involved in superinfection exclusion, persistently CSFV- or LindaV-infected cells were challenged with different pestiviruses. Persistently LindaV-infected cells were significantly more susceptible to cpBVDV infection than persistently CSFV-infected cells, whilst the other pestiviruses tested were not or only hardly able to infect the persistently infected cells. These results provide evidence of a pestivirus species-specific effect on ADAM17 levels and hints at the possibility of its involvement in superinfection exclusion.
Keywords: ADAM17; maturation; pestivirus; receptor downregulation.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Exclusion of Superinfection or Enhancement of Superinfection in Pestiviruses-APPV Infection Is Not Dependent on ADAM17.Viruses. 2024 Nov 26;16(12):1834. doi: 10.3390/v16121834. Viruses. 2024. PMID: 39772144 Free PMC article.
-
Porcine Complement Regulatory Protein CD46 Is a Major Receptor for Atypical Porcine Pestivirus but Not for Classical Swine Fever Virus.J Virol. 2021 Apr 12;95(9):e02186-20. doi: 10.1128/JVI.02186-20. Print 2021 Apr 12. J Virol. 2021. PMID: 33568504 Free PMC article.
-
Inhibition of pestivirus infection in cell culture by envelope proteins E(rns) and E2 of classical swine fever virus: E(rns) and E2 interact with different receptors.J Gen Virol. 1997 Nov;78 ( Pt 11):2779-87. doi: 10.1099/0022-1317-78-11-2779. J Gen Virol. 1997. PMID: 9367363
-
Different Types of Vaccines against Pestiviral Infections: "Barriers" for "Pestis".Viruses. 2022 Dec 20;15(1):2. doi: 10.3390/v15010002. Viruses. 2022. PMID: 36680043 Free PMC article. Review.
-
Structures and Functions of Pestivirus Glycoproteins: Not Simply Surface Matters.Viruses. 2015 Jun 29;7(7):3506-29. doi: 10.3390/v7072783. Viruses. 2015. PMID: 26131960 Free PMC article. Review.
Cited by
-
Exclusion of Superinfection or Enhancement of Superinfection in Pestiviruses-APPV Infection Is Not Dependent on ADAM17.Viruses. 2024 Nov 26;16(12):1834. doi: 10.3390/v16121834. Viruses. 2024. PMID: 39772144 Free PMC article.
References
-
- Jo W.K., van Elk C., van de Bildt M., van Run P., Petry M., Jesse S.T., Jung K., Ludlow M., Kuiken T., Osterhaus A. An Evolutionary Divergent Pestivirus Lacking the Npro Gene Systemically Infects a Whale Species. Emerg. Microbes Infect. 2019;8:1383–1392. doi: 10.1080/22221751.2019.1664940. - DOI - PMC - PubMed
-
- Shi Y.-N., Li L.-M., Zhou J.-B., Hua Y., Zeng Z.-L., Yu Y.-P., Liu P., Yuan Z.-G., Chen J.-P. Detection of a Novel Pestivirus Strain in Java Ticks (Amblyomma Javanense) and the Hosts Malayan Pangolin (Manis Javanica) and Chinese Pangolin (Manis Pentadactyla) Front. Microbiol. 2022;13:988730. doi: 10.3389/fmicb.2022.988730. - DOI - PMC - PubMed
-
- Wu Z., Liu B., Du J., Zhang J., Lu L., Zhu G., Han Y., Su H., Yang L., Zhang S., et al. Discovery of Diverse Rodent and Bat Pestiviruses with Distinct Genomic and Phylogenetic Characteristics in Several Chinese Provinces. Front. Microbiol. 2018;9:2562. doi: 10.3389/fmicb.2018.02562. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
