

Atypical rhizobia trigger nodulation and pathogenesis on the same legume hosts
- PMID: 39461961
- PMCID: PMC11513132
- DOI: 10.1038/s41467-024-53388-x
Atypical rhizobia trigger nodulation and pathogenesis on the same legume hosts
Abstract
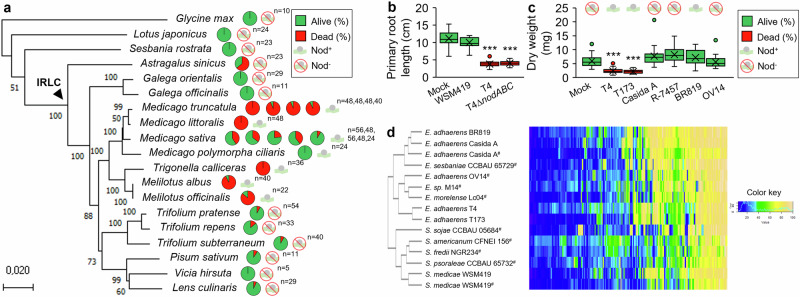
The emergence of commensalism and mutualism often derives from ancestral parasitism. However, in the case of rhizobium-legume interactions, bacterial strains displaying both pathogenic and nodulation features on a single host have not been described yet. Here, we isolated such a bacterium from Medicago nodules. On the same plant genotypes, the T4 strain can induce ineffective nodules in a highly competitive way and behave as a harsh parasite triggering plant death. The T4 strain presents this dual ability on multiple legume species of the Inverted Repeat-Lacking Clade, the output of the interaction relying on the developmental stage of the plant. Genomic and phenotypic clustering analysis show that T4 belongs to the nonsymbiotic Ensifer adhaerens group and clusters together with T173, another strain harboring this dual ability. In this work, we identify a bacterial clade that includes rhizobial strains displaying both pathogenic and nodulating abilities on a single legume host.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests
Figures
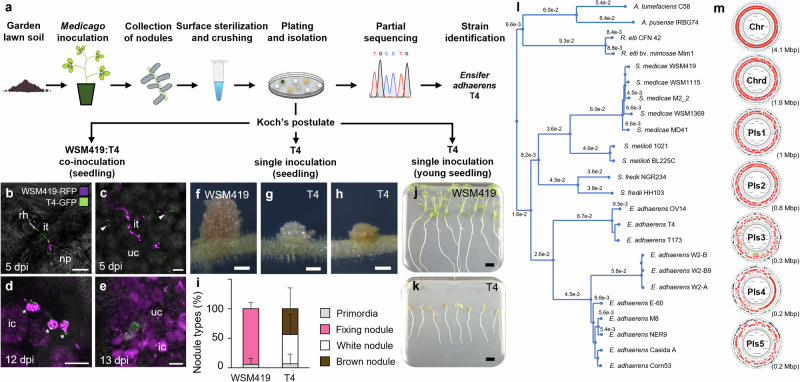
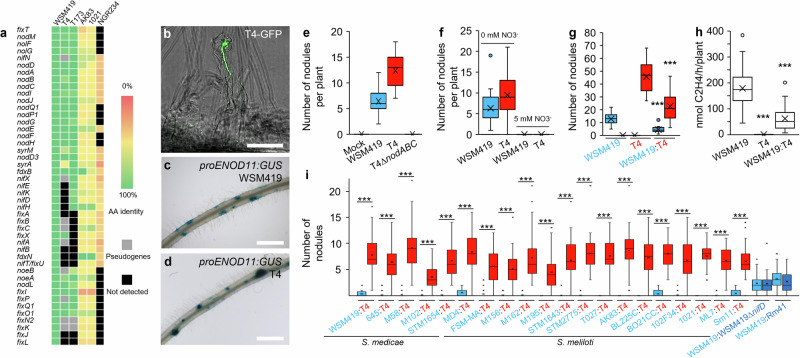
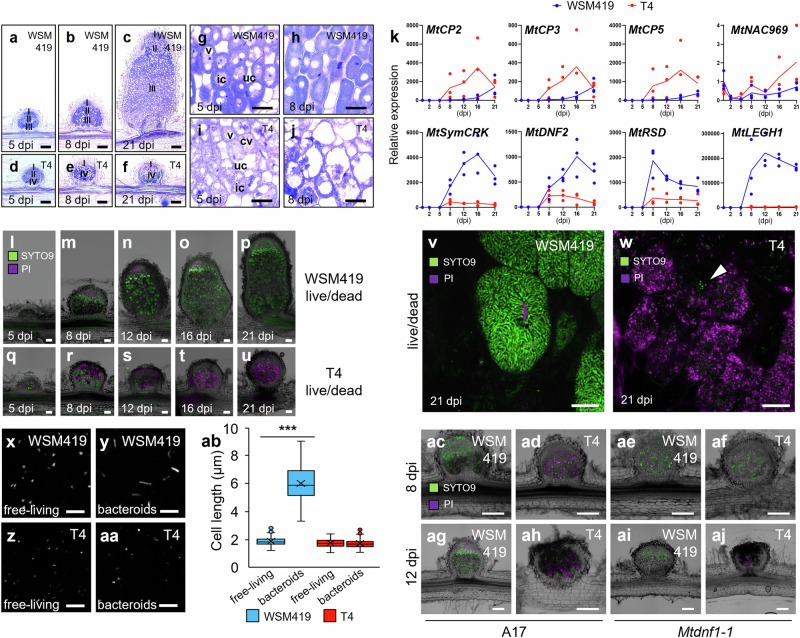
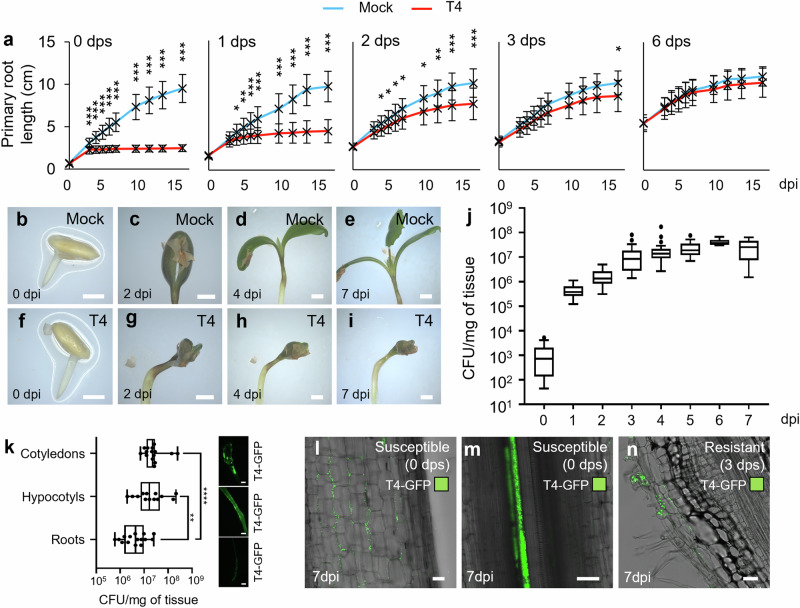
References
-
- Liang, Y. et al. Nonlegumes respond to rhizobial Nod factors by suppressing the innate immune response. Science341, 1384–1387 (2013). - PubMed
Publication types
MeSH terms
Associated data
Grants and funding
- ANR-10-LABX-41/Agence Nationale de la Recherche (French National Research Agency)
- ANR-11-IDEX-0002-02/Agence Nationale de la Recherche (French National Research Agency)
- ANR-21-CE20-0016-01-PATHOSYM/Agence Nationale de la Recherche (French National Research Agency)
- ANR-10-LABX-0040-SPS/Agence Nationale de la Recherche (French National Research Agency)
- EUR SPS-GSR/Agence Nationale de la Recherche (French National Research Agency)
