

The Tmem16a chloride channel is required for mucin maturation after secretion from goblet-like cells in the Xenopus tropicalis tadpole skin
- PMID: 39461969
- PMCID: PMC11514049
- DOI: 10.1038/s41598-024-76482-y
The Tmem16a chloride channel is required for mucin maturation after secretion from goblet-like cells in the Xenopus tropicalis tadpole skin
Abstract
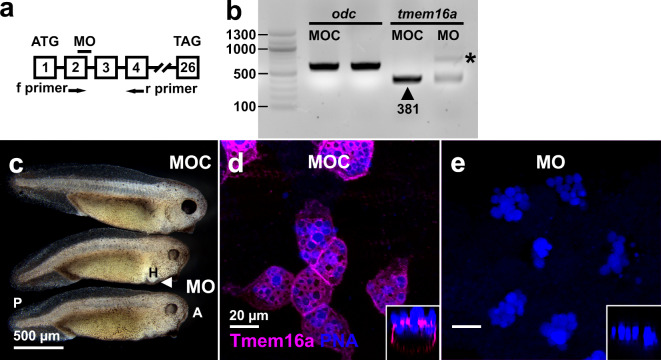
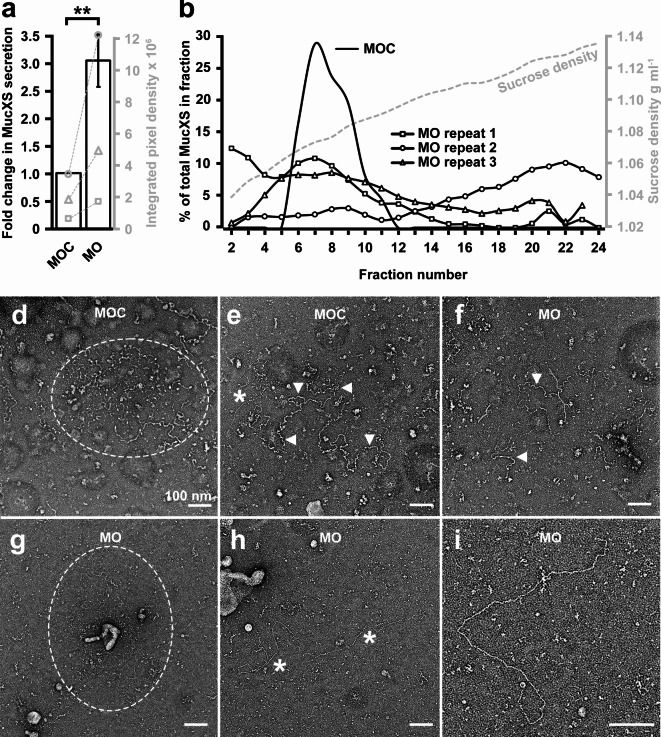
The TMEM16A chloride channel is proposed as a therapeutic target in cystic fibrosis, where activation of this ion channel might restore airway surface hydration and mitigate respiratory symptoms. While TMEM16A is associated with increased mucin production under stimulated or pro-inflammatory conditions, its role in baseline mucin production, secretion and/or maturation is less well understood. Here, we use the Xenopus tadpole skin mucociliary surface as a model of human upper airway epithelium to study Tmem16a function in mucus production. We found that Xenopus tropicalis Tmem16a is present at the apical membrane surface of tadpole skin small secretory cells that express canonical markers of mammalian "goblet cells" such as Foxa1 and spdef. X. tropicalis Tmem16a functions as a voltage-gated, calcium-activated chloride channel when transfected into mammalian cells in culture. Depletion of Tmem16a from the tadpole skin results in dysregulated mucin maturation post-secretion, with secreted mucins having a disrupted molecular size distribution and altered morphology assessed by sucrose gradient centrifugation and electron microscopy, respectively. Our results show that in the Xenopus tadpole skin, Tmem16a is necessary for normal mucus barrier formation and demonstrate the utility of this model system to discover new biology relevant to human mucosal biology in health and disease.
Keywords: Xenopus tropicalis; Ion channel; Mucin; Mucus; TMEM16A.
© 2024. The Author(s).
Conflict of interest statement
HD and MG are employees and stockholders of Enterprise Therapeutics, which has previously developed TMEM16A modulators. ED, ENH, SL, RC, Ch, PM, RKG, ISR and DJT declare no competing interests.
Figures
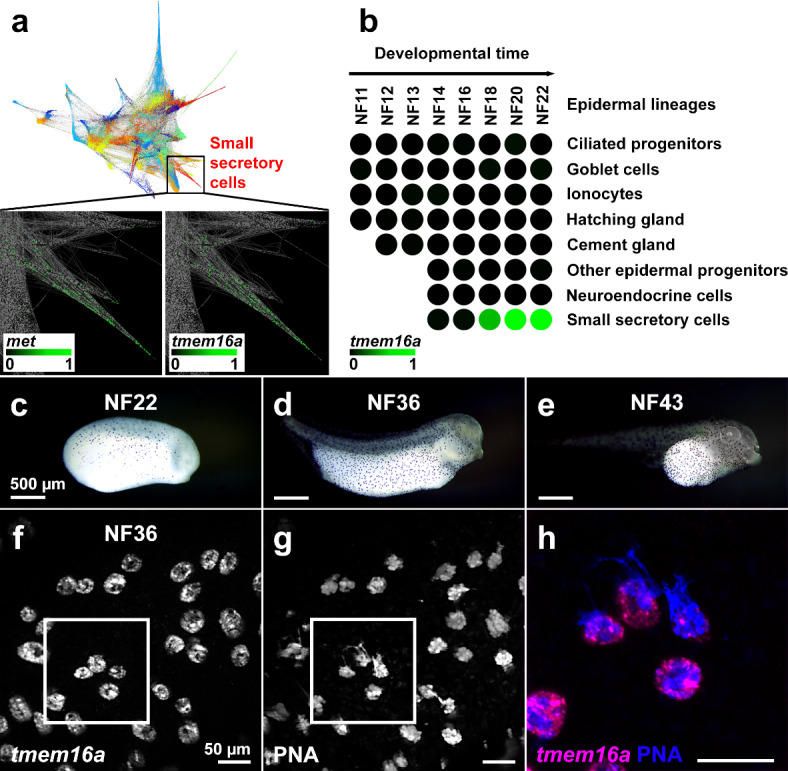
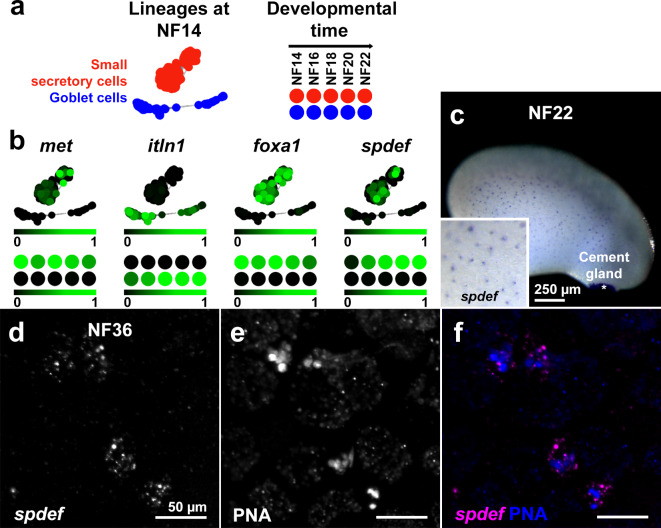
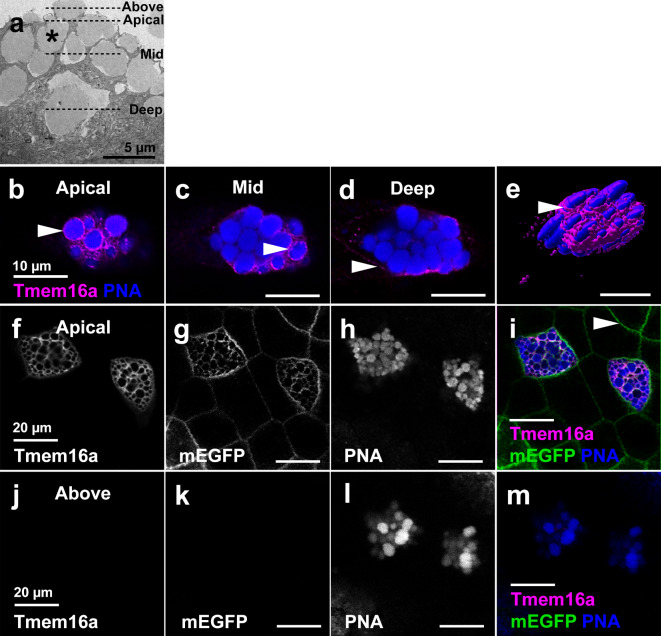
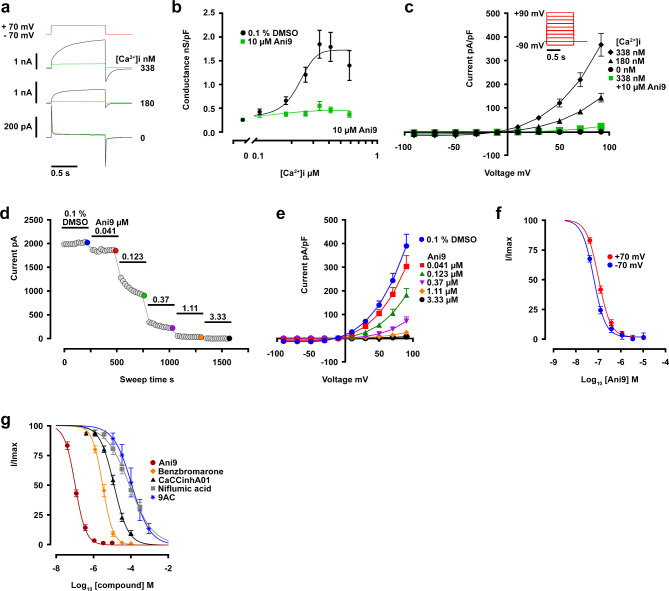
References
-
- Zhou-Suckow, Z., Duerr, J., Hagner, M. & Mall, M. A. Airway mucus, inflammation and remodeling: Emerging links in the pathogenesis of chronic lung diseases. Cell. Tissue Res.367, 537–550 (2017). - PubMed
-
- Kurbatova, P. et al. Model of mucociliary clearance in cystic fibrosis lungs. J. Theor. Biol.372, 81–88 (2015). - PubMed
-
- Rommens, J. M. et al. Identification of the cystic fibrosis gene: Chromosome walking and jumping. Science245, 1059–1065 (1989). - PubMed
MeSH terms
Substances
Grants and funding
- NC/S001034/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- NC/S001034/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- NC/S001034/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- NC/S001034/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- NC/S001034/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- IAA 403/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/W006456/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/W006456/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/W006456/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/W006456/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
