

C G composition in transposon-derived genes is increased in FXD with perturbed immune system
- PMID: 39465205
- PMCID: PMC11500580
- DOI: 10.1093/narmme/ugae015
C G composition in transposon-derived genes is increased in FXD with perturbed immune system
Abstract
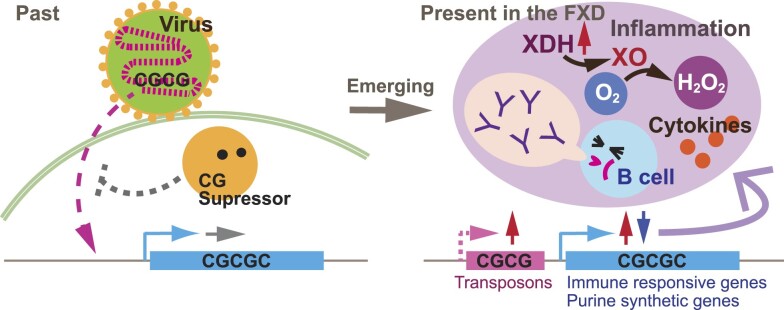
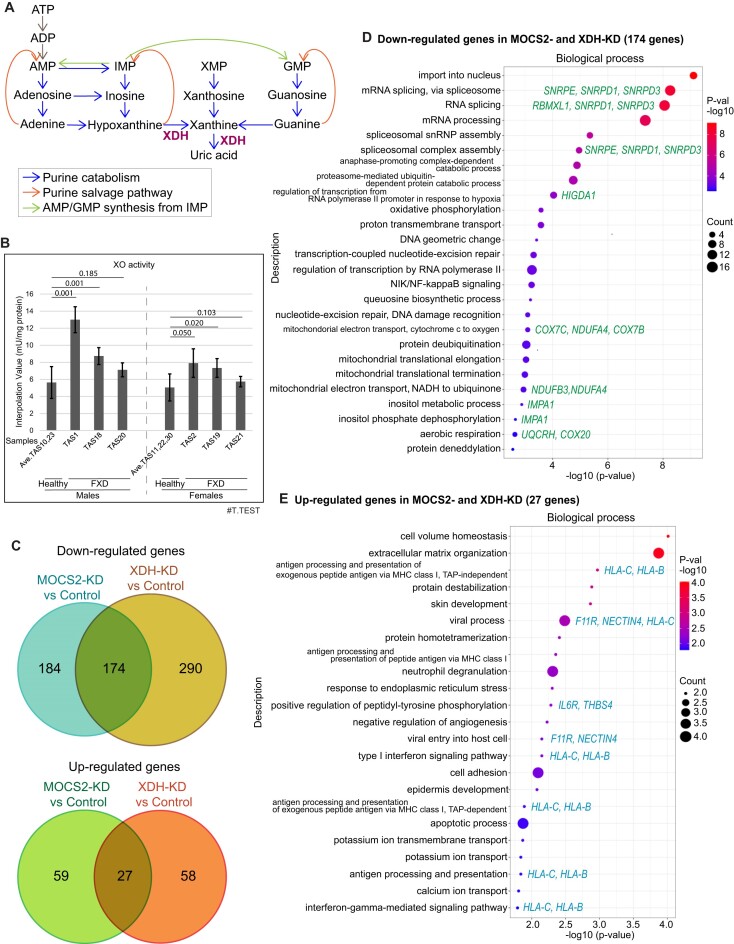
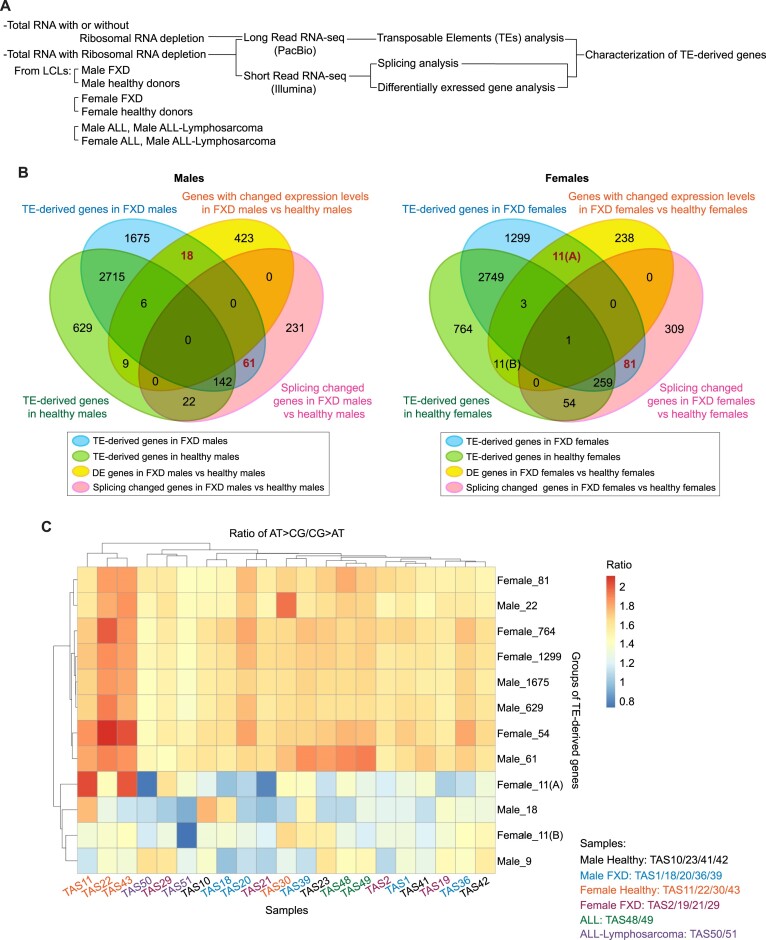
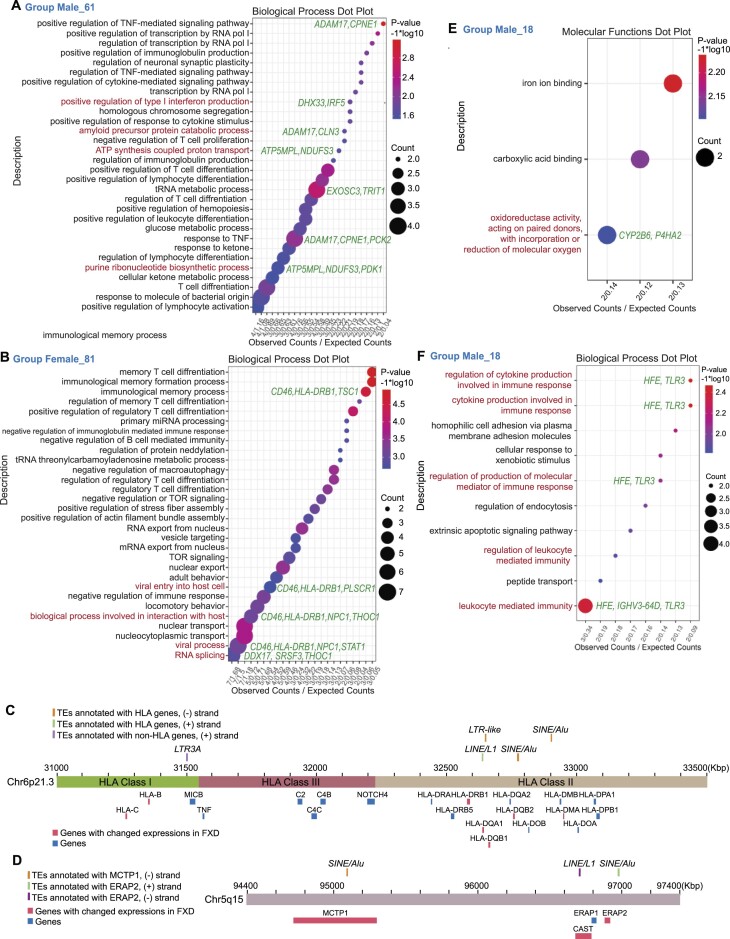
Increasing incidence of Fragile X disorders (FXD) and of immune-mediated disorders in FXD suggests that additional factors besides FMR1 mutations contribute to the pathogenesis. Here, we discovered that the expression levels or splicing of specific transposon element (TE)-derived genes, regulating purine metabolism and immune responses against viral infections are altered in FXD. These genes include HLA genes clustered in chr6p21.3 and viral responsive genes in chr5q15. Remarkably, these TE-derived genes contain a low A T/C G suggesting base substitutions of A T to C G. The TE-derived genes with changed expression levels contained a higher content of 5'-CG-3' dinucleotides in FXD compared to healthy donors. This resembles the genomes of some RNA viruses, which maintain high contents of CG dinucleotides to sustain their latent infection exploiting antiviral responses. Thus, past viral infections may have persisted as TEs, provoking immune-mediated disorders in FXD.
© The Author(s) 2024. Published by Oxford University Press on behalf of NAR Molecular Medicine.
Figures

References
-
- Yu K.H., Palmer N., Fox K., Prock L., Mandl K.D., Kohane I.S., Prilutsky D. The phenotypical implications of immune dysregulation in fragile X syndrome. Eur. J. Neurol. 2020; 27:590–593. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous