

Mechanistic basis for the allosteric activation of NADase activity in the Sir2-HerA antiphage defense system
- PMID: 39465277
- PMCID: PMC11514289
- DOI: 10.1038/s41467-024-53614-6
Mechanistic basis for the allosteric activation of NADase activity in the Sir2-HerA antiphage defense system
Abstract
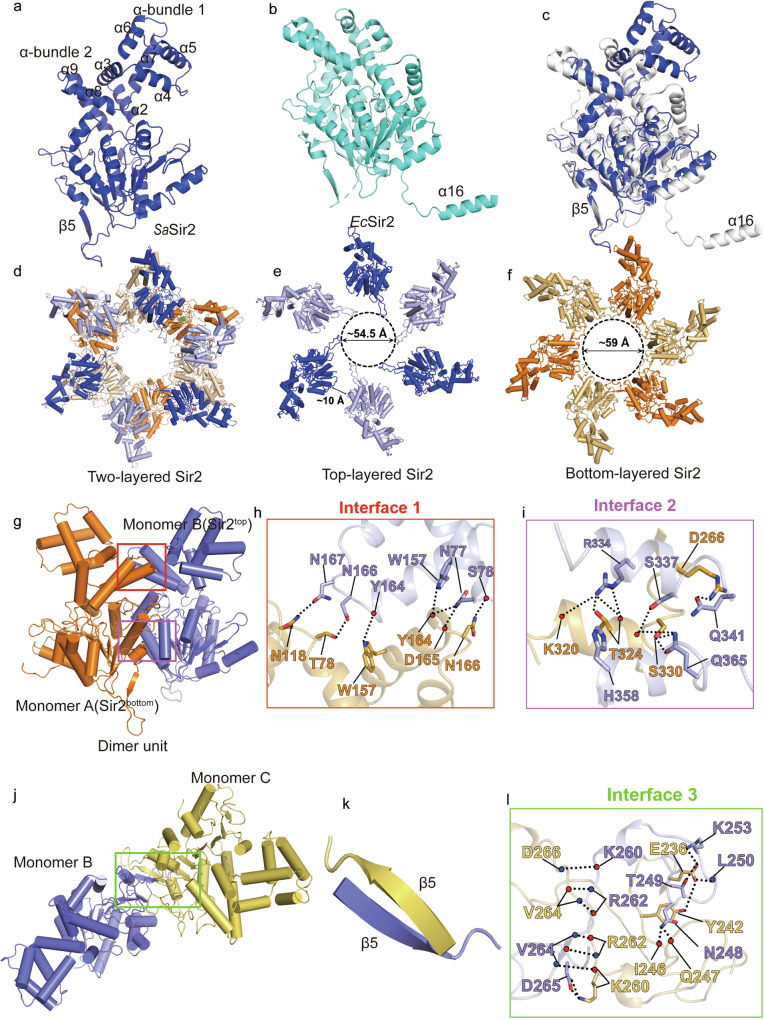
Sir2-HerA is a widely distributed antiphage system composed of a RecA-like ATPase (HerA) and an effector with potential NADase activity (Sir2). Sir2-HerA is believed to provide defense against phage infection in Sir2-dependent NAD+ depletion to arrest the growth of infected cells. However, the detailed mechanism underlying its antiphage activity remains largely unknown. Here, we report functional investigations of Sir2-HerA from Staphylococcus aureus (SaSir2-HerA), unveiling that the NADase function of SaSir2 can be allosterically activated by the binding of SaHerA, which then assembles into a supramolecular complex with NADase activity. By combining the cryo-EM structure of SaSir2-HerA in complex with the NAD+ cleavage product, it is surprisingly observed that Sir2 protomers that interact with HerA are in the activated state, which is due to the opening of the α15-helix covering the active site, allowing NAD+ to access the catalytic pocket for hydrolysis. In brief, our study provides a comprehensive view of an allosteric activation mechanism for Sir2 NADase activity in the Sir2-HerA immune system.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Georjon, H. & Bernheim, A. The highly diverse antiphage defence systems of bacteria. Nat. Rev. Microbiol. 21, 686–700 (2023). - PubMed
-
- Watson, B. N. J., Steens, J. A., Staals, R. H. J., Westra, E. R. & van Houte, S. Coevolution between bacterial CRISPR-Cas systems and their bacteriophages. Cell Host Microbe29, 715–725 (2021). - PubMed
-
- Hille, F. et al. The biology of CRISPR-Cas: backward and forward. Cell172, 1239–1259 (2018). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
