

E2F2 Reprograms Macrophage Function By Modulating Material and Energy Metabolism in the Progression of Metabolic Dysfunction-Associated Steatohepatitis
- PMID: 39465673
- PMCID: PMC11672278
- DOI: 10.1002/advs.202410880
E2F2 Reprograms Macrophage Function By Modulating Material and Energy Metabolism in the Progression of Metabolic Dysfunction-Associated Steatohepatitis
Erratum in
-
Correction to "[E2F2 Reprograms Macrophage Function By Modulating Material and Energy Metabolism in the Progression of Metabolic Dysfunction-Associated Steatohepatitis]".Adv Sci (Weinh). 2025 Apr;12(13):e2502901. doi: 10.1002/advs.202502901. Epub 2025 Mar 7. Adv Sci (Weinh). 2025. PMID: 40052229 Free PMC article. No abstract available.
Abstract
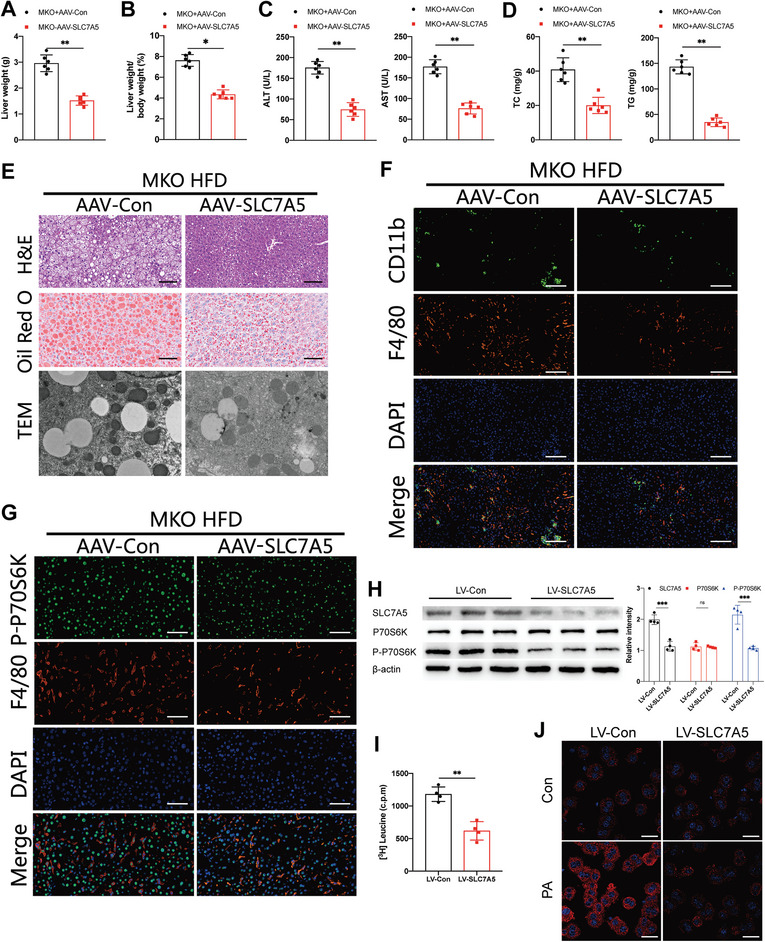
Macrophages are essential for the development of steatosis, hepatic inflammation, and fibrosis in metabolic dysfunction-associated steatohepatitis(MASH). However, the roles of macrophage E2F2 in the progression of MASH have not been elucidated. This study reveals that the expression of macrophage E2F2 is dramatically downregulated in MASH livers from mice and humans, and that this expression is adversely correlated with the severity of the disease. Myeloid-specific E2F2 depletion aggravates intrahepatic inflammation, hepatic stellate cell activation, and hepatocyte lipid accumulation during MASH progression. Mechanistically, E2F2 can inhibit the SLC7A5 transcription directly. E2F2 deficiency upregulates the expression of SLC7A5 to mediate amino acids flux, resulting in enhanced glycolysis, impaired mitochondrial function, and increased macrophages proinflammatory response in a Leu-mTORC1-dependent manner. Moreover, bioinformatics analysis and CUT &Tag assay identify the direct binding of Nrf2 to E2F2 promoter to promote its transcription and nuclear translocation. Genetic or pharmacological activation of Nrf2 effectively activates E2F2 to attenuate the MASH progression. Finally, patients treated with CDK4/6 inhibitors demonstrate reduced E2F2 activity but increased SLC7A5 activity in PBMCs. These findings indicated macrophage E2F2 suppresses MASH progression by reprogramming amino acid metabolism via SLC7A5- Leu-mTORC1 signaling pathway. Activating E2F2 holds promise as a therapeutic strategy for MASH.
Keywords: amino acid transportation; glycolysis; macrophage; metabolic dysfunction‐associated steatohepatitis; slc7a5.
© 2024 The Author(s). Advanced Science published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Rinella M. E., Lazarus J. V., Ratziu V., Francque S. M., Sanyal A. J., Kanwal F., Romero D., Abdelmalek M. F., Anstee Q. M., Arab J. P., Arrese M., Bataller R., Beuers U., Boursier J., Bugianesi E., Byrne C. D., Castro Narro G. E., Chowdhury A., Cortez‐Pinto H., Cryer D. R., Cusi K., El‐Kassas M., Klein S., Eskridge W., Fan J., Gawrieh S., Guy C. D., Harrison S. A., Kim S. U. p, Koot B. G., et al., J Hepatol 2023, 79, 1542. - PubMed
-
- Younossi Z., Anstee Q. M., Marietti M., Hardy T., Henry L., Eslam M., George J., Bugianesi E., Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11. - PubMed
-
- Llovet J. M., Zucman‐Rossi J., Pikarsky E., Sangro B., Schwartz M., Sherman M., Gores G., Nat. Rev. Dis. Primers 2016, 2, 16018. - PubMed
-
- Tincopa M. A., Anstee Q. M., Loomba R., Cell Metab. 2024, 36, 912. - PubMed
-
- Widjaja A. A., Singh B. K., Adami E., Viswanathan S., Dong J., D'Agostino G. A., Ng B., Lim W. W., Tan J., Paleja B. S., Tripathi M., Lim S. Y., Shekeran S. G., Chothani S. P., Rabes A., Sombetzki M., Bruinstroop E., Min L. P., Sinha R. A., Albani S., Yen P. M., Schafer S., Cook S. A., Gastroenterology 2019, 157, 777. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources