

A novel 4-aminoquinoline chemotype with multistage antimalarial activity and lack of cross-resistance with PfCRT and PfMDR1 mutants
- PMID: 39471233
- PMCID: PMC11521309
- DOI: 10.1371/journal.ppat.1012627
A novel 4-aminoquinoline chemotype with multistage antimalarial activity and lack of cross-resistance with PfCRT and PfMDR1 mutants
Abstract
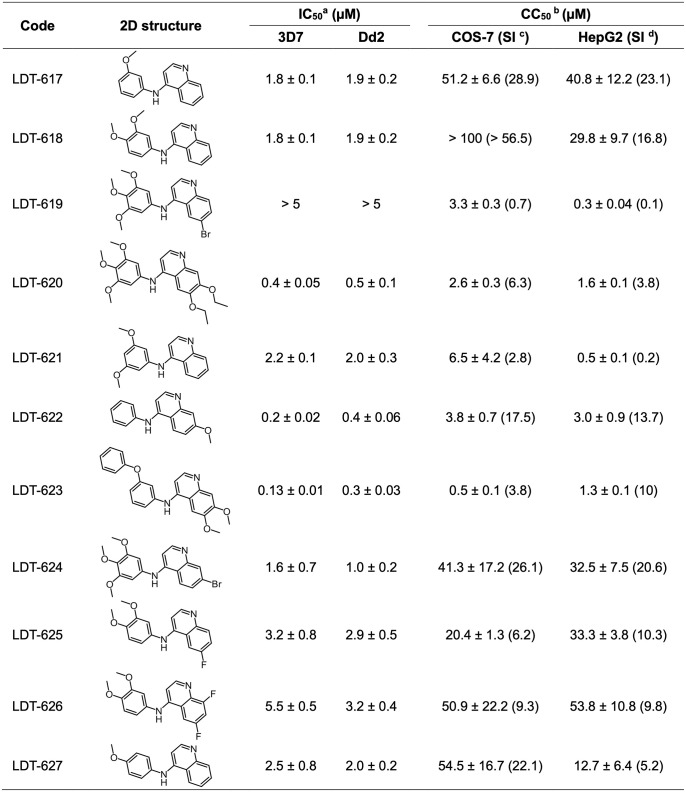
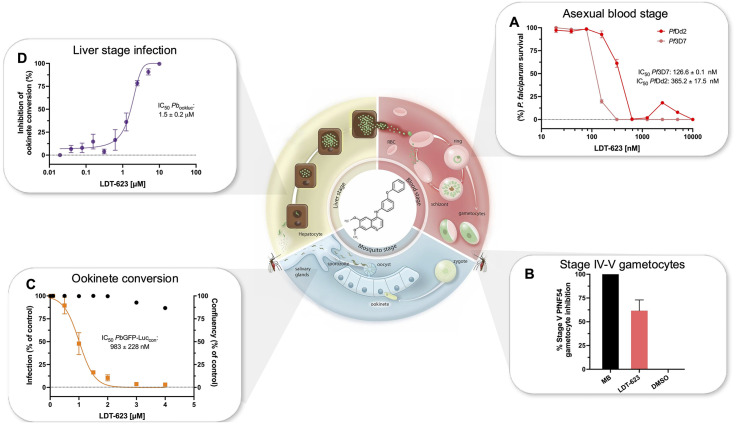
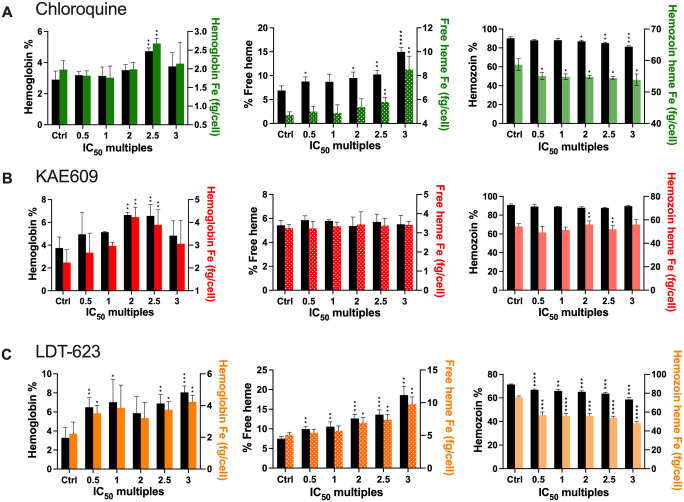
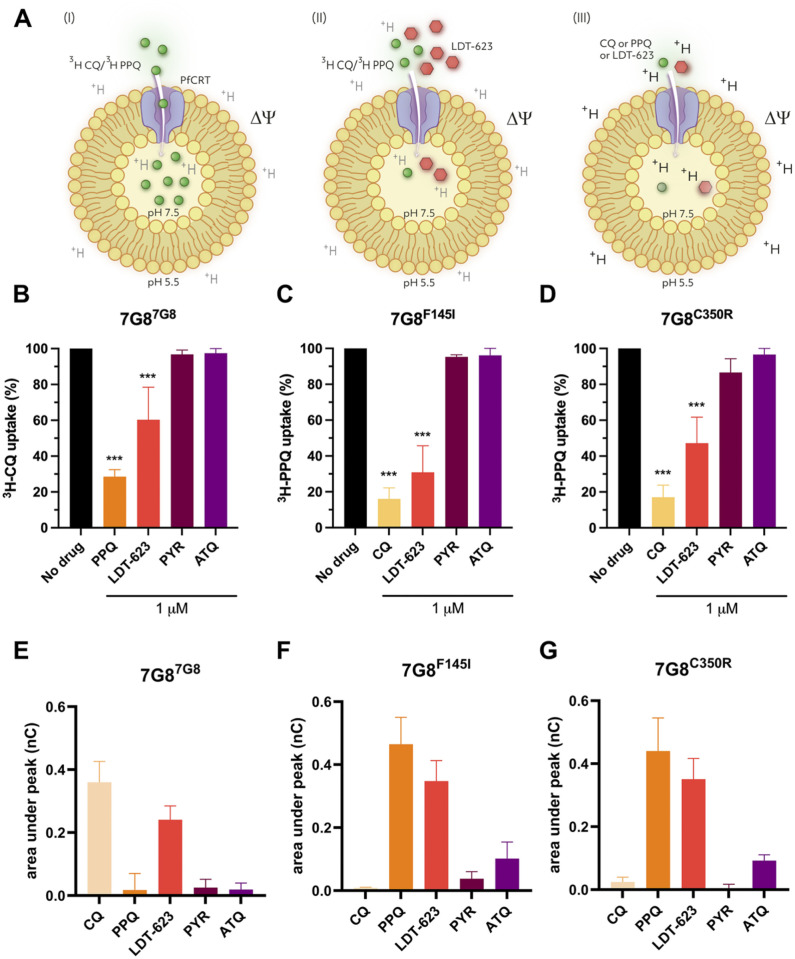
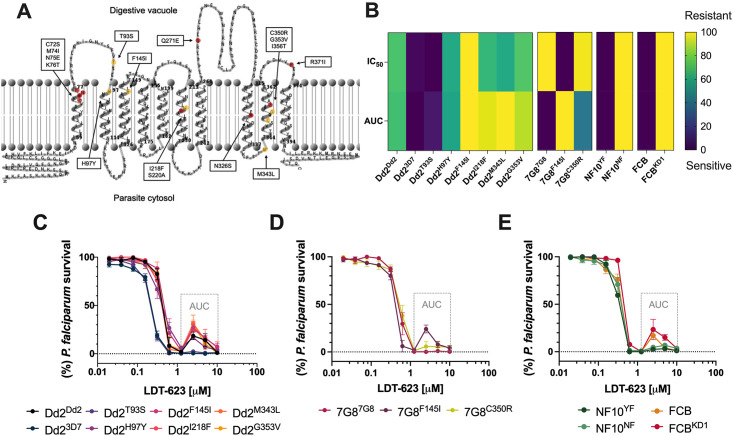
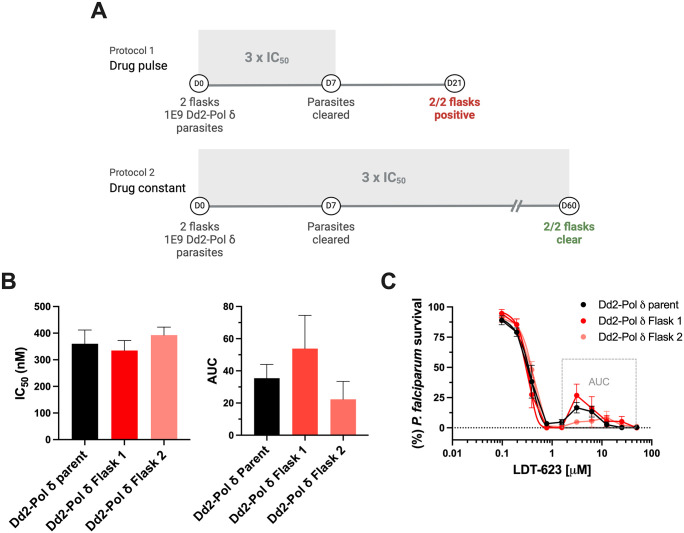
Artemisinin-based combination therapy (ACT) is the mainstay of effective treatment of Plasmodium falciparum malaria. However, the long-term utility of ACTs is imperiled by widespread partial artemisinin resistance in Southeast Asia and its recent emergence in parts of East Africa. This underscores the need to identify chemotypes with new modes of action (MoAs) to circumvent resistance to ACTs. In this study, we characterized the asexual blood stage antiplasmodial activity and resistance mechanisms of LDT-623, a 4-aminoquinoline (4-AQ). We also detected LDT-623 activity against multiple stages (liver schizonts, stage IV-V gametocytes, and ookinetes) of Plasmodium's life cycle, a feature unlike other 4-AQs such as chloroquine (CQ) and piperaquine (PPQ). Using heme fractionation profiling and drug uptake studies in PfCRT-containing proteoliposomes, we observed inhibition of hemozoin formation and PfCRT-mediated transport, which constitute characteristic features of 4-AQs' MoA. We also found minimal cross-resistance to LDT-623 in a panel of mutant pfcrt or pfmdr1 lines, but not the PfCRT F145I mutant that is highly resistant to PPQ resistance yet is very unfit. No P. falciparum parasites were recovered in an in vitro resistance selection study, suggesting a high barrier for resistance to emerge. Finally, a competitive growth assay comprising >50 barcoded parasite lines with mutated resistance mediators or major drug targets found no evidence of cross-resistance. Our findings support further exploration of this promising 4-AQ.
Copyright: © 2024 Ferreira et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- World Health Organization. World Malaria Report 2023. 2023.
-
- van der Pluijm RW, Imwong M, Chau NH, Hoa NT, Thuy-Nhien NT, Thanh NV, et al.. Determinants of dihydroartemisinin-piperaquine treatment failure in Plasmodium falciparum malaria in Cambodia, Thailand, and Vietnam: a prospective clinical, pharmacological, and genetic study. Lancet Infect Dis. 2019;19: 952–61. doi: 10.1016/S1473-3099(19)30391-3 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
