

SERRATE drives phase separation behaviours to regulate m6A modification and miRNA biogenesis
- PMID: 39472512
- PMCID: PMC11631688
- DOI: 10.1038/s41556-024-01530-8
SERRATE drives phase separation behaviours to regulate m6A modification and miRNA biogenesis
Abstract
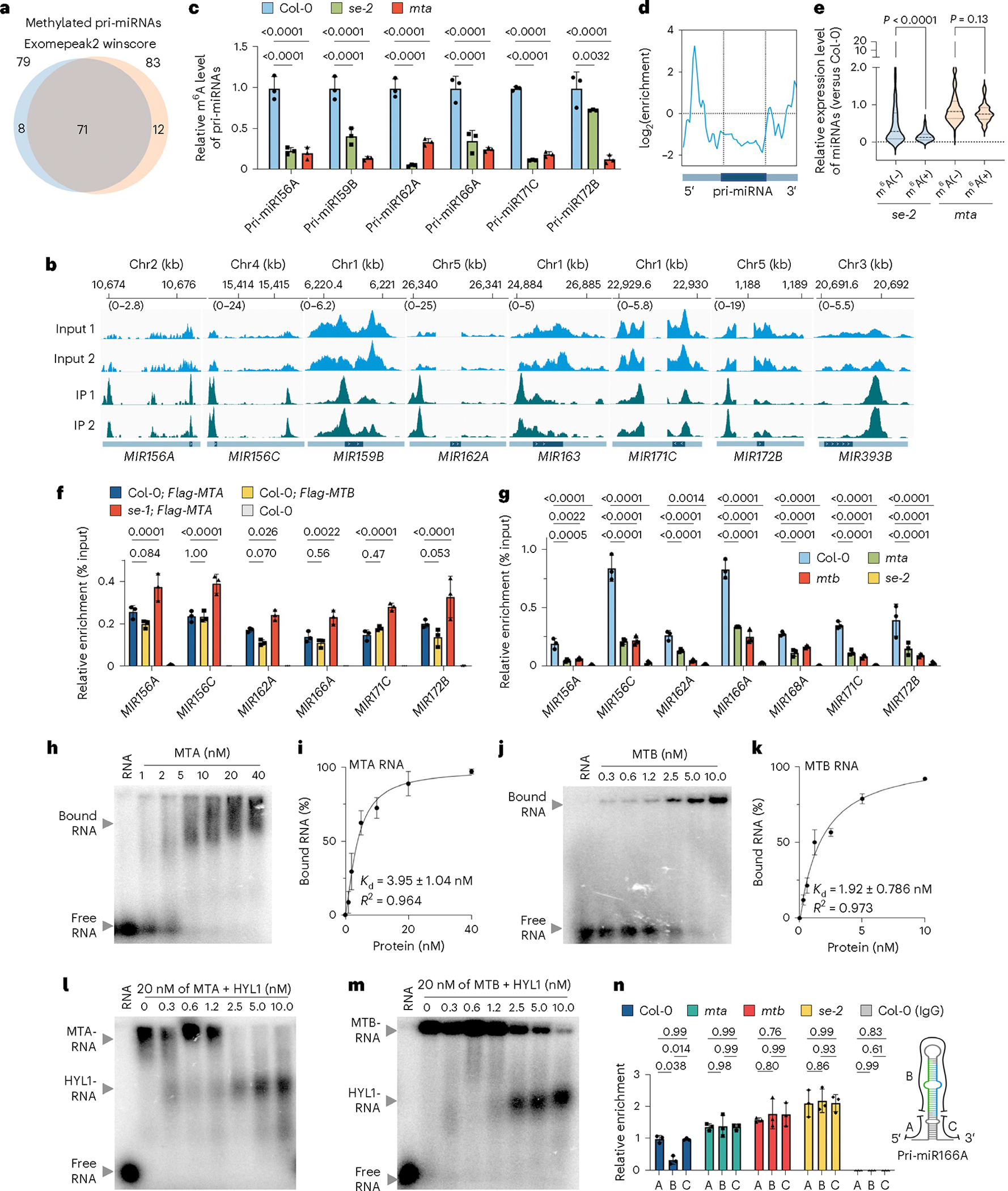
The methyltransferase complex (MTC) deposits N6-adenosine (m6A) onto RNA, whereas the microprocessor produces microRNA. Whether and how these two distinct complexes cross-regulate each other has been poorly studied. Here we report that the MTC subunit B tends to form insoluble condensates with poor activity, with its level monitored by the 20S proteasome. Conversely, the microprocessor component SERRATE (SE) forms liquid-like condensates, which in turn promote the solubility and stability of the MTC subunit B, leading to increased MTC activity. Consistently, the hypomorphic lines expressing SE variants, defective in MTC interaction or liquid-like phase behaviour, exhibit reduced m6A levels. Reciprocally, MTC can recruit the microprocessor to the MIRNA loci, prompting co-transcriptional cleavage of primary miRNA substrates. Additionally, primary miRNA substrates carrying m6A modifications at their single-stranded basal regions are enriched by m6A readers, which retain the microprocessor in the nucleoplasm for continuing processing. This reveals an unappreciated mechanism of phase separation in RNA modification and processing through MTC and microprocessor coordination.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

















References
MeSH terms
Substances
Grants and funding
- 1818082/NSF | BIO | Division of Molecular and Cellular Biosciences (MCB)
- R01 GM127414/GM/NIGMS NIH HHS/United States
- 2139857/NSF | BIO | Division of Molecular and Cellular Biosciences (MCB)
- 31771349/National Natural Science Foundation of China (National Science Foundation of China)
- GM127414/Foundation for the National Institutes of Health (Foundation for the National Institutes of Health, Inc.)
- R35GM151976/Foundation for the National Institutes of Health (Foundation for the National Institutes of Health, Inc.)
- R35 GM151976/GM/NIGMS NIH HHS/United States
- A-2177-20230405/Welch Foundation
- 2019QN01N108/Pearl River S and T Nova Program of Guangzhou (Pearl River S & T Nova Program of Guangzhou)
- R01 GM132401/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
