

Pulmonary SARS-CoV-2 infection leads to para-infectious immune activation in the brain
- PMID: 39474424
- PMCID: PMC11519853
- DOI: 10.3389/fimmu.2024.1440324
Pulmonary SARS-CoV-2 infection leads to para-infectious immune activation in the brain
Abstract
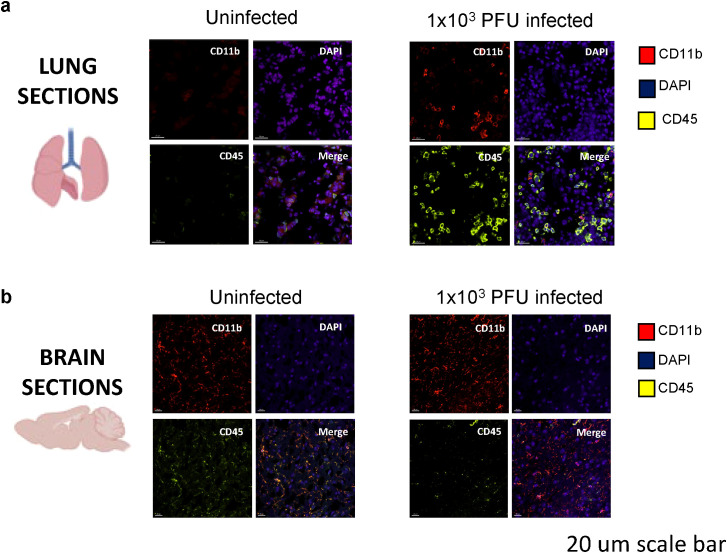
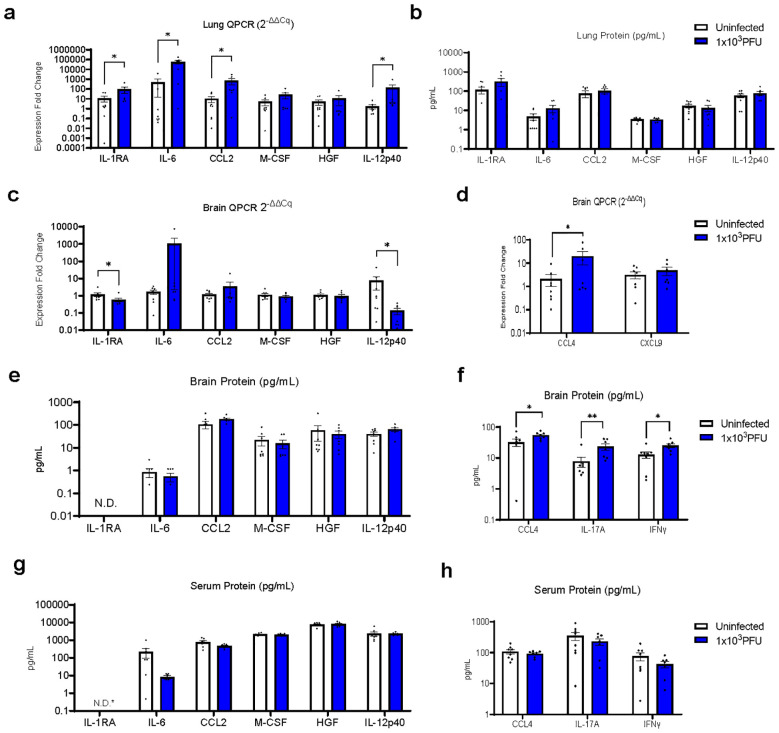
Neurological complications, including encephalopathy and stroke, occur in a significant proportion of COVID-19 cases but viral protein is seldom detected in the brain parenchyma. To model this situation, we developed a novel low-inoculum K18-hACE2 mouse model of SARS-CoV-2 infection during which active viral replication was consistently seen in mouse lungs but not in the brain. We found that several mediators previously associated with encephalopathy in clinical samples were upregulated in the lung, including CCL2, and IL-6. In addition, several inflammatory mediations, including CCL4, IFNγ, IL-17A, were upregulated in the brain, associated with microglial reactivity. Parallel in vitro experiments demonstrated that the filtered supernatant from SARS-CoV-2 virion exposed brain endothelial cells induced activation of uninfected microglia. This model successfully recreates SARS-CoV-2 virus-associated para-infectious brain inflammation which can be used to study the pathophysiology of the neurological complications and the identification of potential immune targets for treatment.
Keywords: SARS-CoV-2; immunology; microglia; neurology; virology.
Copyright © 2024 Dunai, Hetherington, Boardman, Clark, Sharma, Subramaniam, Tharmaratnam, Needham, Williams, Huang, Wood, Collie, Fower, Fox, Ellul, Held, Egbe, Griffiths, Solomon, Breen, Kipar, Cavanagh, Irani, Vincent, Stewart, Taams, Menon and Michael.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous